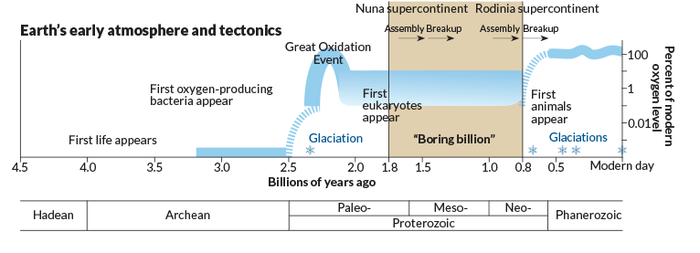
Boring Billion
The Boring Billion refers to the approximately one billion-year period between 1.8 and 0.8 Ga in Earth's history that is characterized by environmental, evolutionary, and lithospheric stability.[1] It has also been termed the "Barren Billion",[2] "Dullest Time on Earth"[3] and "Earth’s Middle Ages".[1] In the time leading up to the Boring Billion, Earth experienced multiple widespread glaciations, the origin of prokaryotic life, the introduction of oxygen into the atmosphere with the evolution of cyanobacteria, addition of UV-blocking ozone to the atmosphere, and the oxidizing of iron in the oceans.[4] After the Boring Billion, the atmosphere again underwent rapid changes as atmospheric oxygen rose to approximately modern levels, most major animal phyla evolved during the Cambrian explosion, and large animals appeared in the oceans.[4] The Boring Billion was thus termed boring because unlike the rapidly changing environments present on Earth before and after this period, it is characterized by climatic stability, low levels of atmospheric oxygen, lack of biological events, and the absence of extreme changes in the atmospheric and oceanic composition.[5][6][7] Stability during the Boring Billion may be attributed to a relatively stable supercontinent that was initiated by 1.7 Ga and persisted until its breakup around 0.75 Ga.[4] The exact timing and duration of the Boring Billion is not agreed upon by scientists and estimates for the beginning and end of the Boring Billion range between 1.8 and 2.4 Ga for initiation and between 0.5 and 0.8 Ga for termination. The Boring Billion occurred during the Proterozoic Eon.
During the Boring Billion, green and purple photosynthetic bacteria appear to have thrived in an anoxic and sulfidic ocean.[4] This ocean was much less productive than modern oceans, released sulfurous gasses including toxic hydrogen sulfide, and was very limited in nutrients (especially Mo, Fe, N, and P).[8][9] As the Boring Billion progressed, eukaryotic life evolved from a prokaryotic ancestor within this ocean.[10] By the end of the Boring Billion, the first life had appeared on land.[11] Eukaryotes, specifically a proto-lichen, helped end the Boring Billion by causing Earth’s second oxygenation event and the Snowball Earth glaciation that accompanied it.[4]
While the Boring Billion has been termed ‘boring’ due to its relative stability, scientists acknowledge that this name might not accurately describe this time.[5] When plate tectonics, environmental conditions, and evolution of eukaryotic life are examined, this period does not appear boring at all.[5][12] Records of this distant time are sparse, and the apparent stability of this period may be due in part to gaps in the geologic record.[13]
Climatic stability
Recent scientific studies on the Boring Billion have found little evidence of significant climatic variability during this time period.[2][14] Different theories have been developed by individual scientists to explain the inferred climatic stability, while others[5] attribute the lack of evidence to an absence of suitable records. There is currently no generally accepted theory in the scientific community on the cause(s) of the climatic stability during the Boring Billion.
Tectonic constraints
The evolution of Earth’s biosphere, atmosphere and hydrosphere has long been linked to the supercontinent cycle, through which the Earth’s continental crust undergoes a periodic cycle of aggregation and dispersal. The period of the Boring Billion is characterized by the evolution of two different supercontinents: Columbia and Rodinia. The supercontinent Columbia amalgamated between 2.0 to 1.7 Ga and remained as a quasi-integral continental lid until at least 1.3 Ga. Several breakup attempts were unsuccessful. Geological and paleomagnetic evidence suggest that Columbia underwent only minor changes to form the supercontinent Rodinia during the period from 1.1 to 0.9 Ga. Paleogeographic reconstructions suggest that the supercontinent assemblage was located in equatorial and temperate climate zones throughout the intervening time frame; there is little or no evidence for continental fragments in polar latitudes.[15] A consequence of the limited breakup history is the lack of passive margins during the time period from 1.8 to 0.8 Ga.[16] This stable configuration provides the primary constraint on the environmental stasis that characterizes the oceans, atmosphere, and biosphere. The breakup of Rodinia at approximately 0.75 Ga is considered the end of Earth’s Boring Billion.[1] It should be noted that numerous reconstructions of Columbia’s paleogeography exist and consensus has yet to be reached. Evaluating the evidence of paleo poles and continent location becomes increasingly more difficult as deposits get older and relevant rock units show increasing levels of deformation and fragmentation.[5]
The relative scarcity of passive margins during the Boring Billion, which are created during times of continental breakup, is proposed to support the presence of a stable continental configuration enabling environmental and climate stability during this time period.[1][5] However, it is also stated in the literature that in spite of the amalgamation of the supercontinents Columbia and Rodinia, relatively stable climatological conditions during the Boring Billion are consistent with a balance between weathering intensity, mantle temperatures and solar insolation throughout much of this time period.[2]
A mechanism put forward to explain low tectonic variability is related to the temperature of the asthenosphere, which may have been too hot to sustain modern plate tectonics in the early stages of the Earth’s evolution. Instead of vigorous plate recycling at subduction zones, plates were linked together for billions of years until the mantle cooled off enough to further sustain plate motions and the subsequent formation of subduction zones.[17] The onset of this component of plate tectonics may have been aided by the thickening of the crust that, once initiated, caused plate subduction to be anomalously strong.[1] This process occurred around 750 Ma ago, when the crust reached a temperature low enough to initiate and sustain plate tectonics in general and subduction processes in particular.[1]
Cosmic activity
The absence of glaciation could be linked to the intensity of cosmic ray flux, and it is believed that periods of glaciation may be linked to periods with low cosmic ray flux due fluctuations in solar wind variations.[18] Another possible cause is the rate of star formation in the Milky Way. The reduced rate in star formation may be linked to diminished glaciations from 1 to 2 Ga.[19]
Absence of prolonged glaciations
The Boring Billion as a time period is unique in a sense that it seems to lack any significant periods of prolonged glaciations that can be observed in regular periodicity in other parts of Earth's geologic history.
Some researchers[20][21] believe, however, that the proposed lack of an ozone layer, as expected during periods of low concentrations of atmospheric oxygen, in combination with lower solar intensity[22] during that time period should have precluded the absence of glaciation without an intense greenhouse effect.
Instead, the absence of a glacial record may be a function of data infidelity rather than a real feature of geologic time. Evidence for this are old glacially cut channels, which were found in the Kimberley district of Western Australia that have been dated to 1.8 Ga.[13]
Low oxygen levels
Earth's geologic record indicates two events associated with significant increases in oxygen levels on Earth, with one occurring between 2.4 to 2.1 Ga, known as the Great Oxygenation Event (GOE), and the second occurring an approximate 0.8 Ga.[23]
The period of time between these two oxygenations is thought to have been characterized by low levels of atmospheric oxygen (O2). Researchers postulate this widespread anoxia was far greater than the anoxic deep oceans seen today (less than 1% of deep ocean). The cause and extent of the oxidation events and the subsequent billion years in between are subject to active debate. Although it was previously believed that atmospheric oxygen levels increased gradually through this period, new research has suggested that after the GOE - in which O2 levels spiked over 1,000 fold - oxygen levels dropped significantly and remained low (with minor fluctuations) until the second oxygenation event around 0.8 Ga.[7]
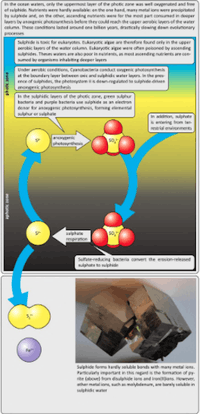
The Lomagundi Excursion Event is considered to be the most prominent carbon isotope event in Earth’s history and can provide evidence for this large increase and subsequent decline in O2. For this event, a release of 10 to 20 times the current atmospheric oxygen is predicted based on levels found in the carbonate δ13C record around the time of the GOE. Suggestions in the literature point to a ‘precipitous’ drop after the Lomagundi Excursion.[24][25][26] The extent to which the GOE served as a ‘Great’ oxygenation event, particularly in regards to marine environments, became a topic of discussion in the scientific community, leading Donald Canfield to develop the idea of a two billion year lag in oxygenation of the oceans. He also proposed that the ocean was characterized high levels of hydrogen sulfide during this time, creating a euxinic environment. His explanation has become known as the Canfield Ocean hypothesis. It is believed that the low and fluctuating oxygen levels that characterized the Boring Billion effectively stalled evolution of complex life for the extent of the Boring Billion. The hypothesis of a predominantly globally euxinic ocean, with low to no oxygen, supports the theory of stalled evolution of complex animals throughout the Boring Billion. Complexity is believed to have evolved at the onset of a second oxygenation event and subsequent oxygenation of the ocean between 600-800 Ba.[27][28]
Canfield ocean hypothesis
Donald Canfield proposed a hypothesis to explain how life persisted in the ocean despite low levels of oxygen.[7] It is believed that during the Boring Billion, the ocean’s surface contained low levels of oxygen as a byproduct of oxygenic photosynthesis from cyanobacteria. Due to increased oxygen levels in the atmosphere, chemical weathering of the continents oxidized the mineral pyrite and transported sulfate (SO42-) to the oceans.[29] Through this process, the ocean became rich in sulfur, and the ocean became a two layer system, consisting of a surface layer, high in oxygen, and a bottom layer, high in sulfur and low in oxygen. Canfield proposed that at the boundary between the two layers, green sulfur bacteria and purple bacteria served to convert the sulfur to hydrogen sulfide (H2S). This process of deep sea sulfide creation led to high amounts of pyrite formation during the Boring Billion, and allowed for the chemical events that enabled the evolution of animals.[30][31]
Johnston ocean hypothesis
Like the Canfield Ocean model, the Johnston hypothesis serves as a potential explanation of causal factors for tempered oxygenic production. The model ultimately claims that sulfide driven anoxic photosynthesis via sulfidic prokaryotes helped sustain a long period of deep ocean anoxia, and more specifically, euxinia. This period ended with the exhaustion of sulfide resources, leading to the coupling of oxygenic production and carbon burial that is characteristic of today’s oceans.[32]
At the first order, Johnson’s take on the Canfield model is to recognize that maintaining the anoxygenic conditions requisite for a billion years would be difficult: as organic matter sinks, it decays and consumes oxygen, a process which is exacerbated by anoxygenic photosynthesis. It has been hypothesized that an explanation for this discrepancy could be anoxygenic photosynthesis, a primary production pathway that does not involve oxygen production as a byproduct. In this case, the consumption of oxygen by respiration is not balanced by oxygen generation through primary production. A modern example of such a system is best exemplified in parts of the Black Sea.[7]

Further evidence
Gaps in the evidence from ancient rocks have failed to explain the discrepancy between Earth’s atmospheric and oceanic oxygen levels. By tracing the movement of chromium isotopes from land to ocean, a process that is sensitive to atmospheric oxygen levels, the earliest signs of significant chromium oxidation have been found. This suggests levels of atmospheric oxygen to have significantly increased around 800 million years ago. Small amounts of oxygen can convert insoluble chromium, a constituent of the continental crust to a soluble form which is then transported to the ocean and deposited in ironstone. By studying the chromium isotope composition in ancient ironstones, measuring the ratio of heavier 53Cr (which is more sensitive to oxidation) to the lighter 52Cr, researchers can determine ancient oxygen levels. This research revealed very low oxygen levels during the Boring Billion, which are below 0.1% of present atmospheric levels.[27]
Rather than iron oxidation involving iron (III), pyrite formation (FeS2) involves the uptake of iron (II) during euxinic conditions. A research team from the University of Tasmania, under the geologist Ross Large, measured levels of pyrite in ancient seafloor rocks to show that after the initial burst of oxygen 2.4-2.1 Ga there was a decline in oxygen levels until the second oxidation event around 800-600 Ma.[28]
Earth's first eukaryotes
In comparison to the great increase in diversity during the Cambrian explosion, the evolution of life during the Boring Billion was relatively slow.[33] However, this period did see the occurrence of several major events in the history of evolution, specifically the development of eukaryotic cells, the evolution of multicellularity and the evolution of sexual reproduction.[4] Several steps were required in the evolution of eukaryotes from a prokaryotic ancestor including a reorganization of DNA within cells and the acquisition of organelles.[4]
The first eukaryotes originated sometime between 2.3 and 1.1 billion years ago, around the beginning of the Boring Billion.[4] The precise onset of eukaryotic evolution is hard to determine, but evidence for the presence of eukaryotic cells during this period can be found in the fossil record and molecular fossils.[4] After eukaryotic cells evolved, multicellularity appeared.[4] By 1.8 Ga the first living things on planet Earth that were big enough to see with the naked eye had appeared.[4] By the end of the Boring Billion, multicellular algae, fungi, and animals inhabited Earth.[4] Almost all of this life was restricted to Earth's oceans, but near the end of the Boring Billion the first terrestrial life, a proto-lichen, colonized land.[4]
Steps required for eukaryotic evolution
There is no agreed upon theory for the evolutionary steps that led to the appearance of the first eukaryotes, including the origin of the nucleus, cytoskeleton, and organelles that distinguish eukaryotes from prokaryotes (See "Hypotheses for the origin of eukaryotes" within the Eukaryote page). There are several changes that must have occurred as eukaryotic life evolved: an increase in genetic complexity, the development of an advanced cytoskeleton, a loss of the rigid bacterial outer wall, and the acquisition of membrane-bound organelles within cells.[4]
DNA reorganization
Eukaryotic cells are generally much more complex than prokaryotic cells. When eukaryotic life evolved, an increase in genetic material was necessary to enable this complexity. With an increase in the amount of genetic material inside cells as eukaryotes were evolving, the evolution of entirely new structures and mechanisms of gene copying was required, leading to the development of the eukaryotic cell cycle.[4]
Histones likely appeared during the Boring Billion to help organize the increasing amount of DNA in ancestral eukaryotic cells.[4] DNA was packed by histones into the first chromosomes. A system of more rapid eukaryotic DNA replication in which DNA was replicated from multiple replication origins was developed so that cells containing much more genetic material would still be able to reproduce quickly.[4]
Advanced cytoskeleton
The development of an advanced cytoskeleton was necessary to organize the chromosomes during asexual cell replication.[4] After the loss of the rigid bacterial outer wall, the advanced cytoskeleton allowed the cells to flow and move.[4] The loss of the rigid bacterial cell wall, which helped prevent water from entering the cell and diluting the cytoplasm, required ancestral eukaryotes to develop pumping mechanisms to conteract osmosis.[4] Once the advanced cytoskeleton was in place and cell wall removed, ancestral eukaryotic cells had the ability to flow around and engulf other cells using the process of phagocytosis, which is a more efficient way of obtaining nutrients than the system of external absorption used by prokaryotes.[4]
Acquisition of organelles (first endosymbiosis)
The ability to ingest other cells enabled ancestral eukaryotes to build a much more complex cell.[4] Once the capacity for consuming other cells was in place, early eukaryotes began to acquire internal membrane-bound organelles that included organelles that were once free-living prokaryotes.[4] Based on phylogenetic and structural evidence, it is generally accepted that mitochondria were originally derived from a free-living a-proteobacterium.[10] The bacterium was engulfed by an ancient cell, likely an Archaea, and entered into the first endosymbiotic relationship with its new host.[10]
Most models have explained the origin of the endosymbiosis in terms of the advantage gained by the host cell due to the greater energy production available through aerobic respiration (32 net ATP vs. 2 net for glycolysis). However, this hypothesis has several limitations. No known organism exports ATP to its environment, making it unlikely that a bacterium would begin doing so to its host.[34] In addition, some eukaryotes possess mitochondria-derived organelles called hydrogenosomes which perform a type of anaerobic respiration, suggesting an adaptation to an anoxic environment.[34] An alternative explanation known as the Hydrogen Hypothesis suggests that the unique conditions of the Canfield Ocean may have exerted a positive selective pressure on mitochondria for reasons unrelated to aerobic respiration. According to the Hydrogen Hypothesis, the original endosymbiotic event was between a hydrogen-producing bacterium and a hydrogen-consuming archaeon.[35] This symbiosis would have been advantageous in the anoxic conditions in the deeper reaches of the oceans at that time and would explain the widespread phylogenetic distribution of anaerobic respiration genes among eukaryotes.[35] One other hypothesis is that a sulfur-metabolizing archaea and the ancestor of mitochondria lived in proximity, each feeding on the other's waste products and deriving energy through the recycling of sulfur.[36] When environmental conditions became harsh for one partner, the energy generation and recycling system became internalized and the archaeon and ancestral mitochondria fused to form one organism.[36]
Chloroplasts are also considered endosymbionts and were likely acquired during the Boring Billion in the same manner as mitochondria.[4] It is generally accepted that chloroplasts are descendants of a free-living oxygen-producing cyanobacteria.[4]
Fossil evidence for eukaryotic evolution
While it is unknown how the first eukaryotes evolved, they first appear in the fossil record by 1.85 Ga.[4]
Acritarchs, whose name means ‘uncertain origin’, are tiny, roughly spherical fossils present from before 2 billion years ago and throughout the Boring Billion that cannot be convincingly assigned to any particular part of the tree of life.[37] One interpretation of acritarchs is that they represent the resting stage, or cysts, of eukaryotic algae.[4] However, since bacteria are also known to cysts and spores, a prokaryote origin can’t be ruled out. It is generally agreed by experts on fossil life that acritarchs from about 1.85 Ga and later represent eukaryotes.[4]
Larger two centimeter, 2.1 billion year old spiral fossils found in a mine in Michigan are grouped into the Grypania, a grouping of spiral fossils previously thought to have arisen 1.4-1.6 billion years ago.[38] Traces of internal detail confirm that the younger Grypania fossils are eukaryotes, but the older spirals lack fine detail and may have been made by filaments of prokaryote cells.[39] The older fossils were possibly also incorrectly dated and have a more accurate age of 1.8-1.9 billion years.[40]
Molecular fossil evidence for eukaryotic evolution
Steranes, the traces of sterols used in construction of the membranes of eukaryotes, have been found in 2.7 billion year old shales, and are evidence that eukaryotes, or at least their metabolic ancestors, were present at that time.[41] While the sterols found in these shales are not made by any known modern bacteria, some bacteria are capable of producing sterols. Therefore, sterane presence is not definite evidence of eukaryotic cells.[42]
Eukaryotic diversification
After the first eukaryotes appeared, the diversification and evolution of eukaryotes proceeded slowly.[4] Environmental conditions, especially the presence of toxic sulfide, lack of oxygen in ocean water, lack of key nutrients and trace metals in ocean water, and/or little tectonic activity, contributed to slow eukaryotic evolution.[1] Low oxygen levels in ocean water during the Boring Billion may have kept eukaryotes in low numbers and prevented the evolution of large energy hungry, fast moving animals that evolved at the end of the Boring Billion,[20] though it has been argued that even with very little oxygen, animals could have emerged and persevered.[43] It is also known that diversification of eukaryotes began well before the onset of global oxygenation of the oceans, leading to the conclusion that anoxic conditions could not be the only cause of slow eukaryotic evolution.[4]
The anoxic and euxinic conditions of the period would also have affected the abundance of certain nutrients and trace metals in the environment. Both anoxic and sulfidic conditions lead to decreased solubility of iron,[44] while sulfidic conditions lead to increased precipitation of the trace metal molybdenum.[45] Both of these metals are essential for the utilization of nitrogen, as they are necessary for the enzymes nitrate reductase, which reduced nitrate and nitrite to ammonia, and nitrogenase, which reduces atmospheric nitrogen to nitrate.[46] As such, life in the Proterozoic would have suffered from a lack of nitrogen, a situation which would have competitively favored prokaryotes over eukaryotes and likely negatively affected eukaryotic evolution.[46]
Despite these environmental constraints, eukaryotes developed multicellularity during the Boring Billion and a major diversification of eukaryotes occurred at the supergroup level between 1.9 and 0.75 Ga.[4] Key biological innovations such as cell differentiation, sexual reproduction, and eukaryotic photosynthesis separated eukaryotes into animals, algae, and fungi.[4]
Multicellularity
The evolution of cell polarity, which allows cells to send a molecular message to one side of the cell but not the other and to interact with nearby cells, allowed the development of organized multicellularity during the Boring Billion.[4] Qingshania, a small spiral coil that appears in the fossil record 1.7 Ga, is the oldest known fossil multicellular eukaryote.[47]
Cellular Differentiation and Sexual Reproduction
The much larger eukaryotic genome with its rapid copying gave eukaryotes the capacity to create many different types of cells and assemble them together into large, complex, multicellular life.[4] By 1.2 Ga, meiosis and sexual reproduction are present in single-celled eukaryotes.[48] Bangiomorpha pubescens, a fossil multicellular red algae from the 1.2 Ga Hunting Formation, is the first known sexually reproducing organism. Fossils show differential spore/gamete formation, which is also evidence of cellular differentiation.[49]
First Algae
The first algae split from ancestral eukaryotes about 1.4 Ga.[4] Molecular clock evidence suggests that red and green algae had separated by 1.2-0.9 Ga.[4]
First Fungi
From molecular clock dating of DNA and fossil evidence, the divergence of animals and fungi occurred between 1.2 and 1.9 billion years ago.[50] 1.43 Ga fossils named Tappania may represent the first fungi.[4] These 0.1 mm fossils of branching irregular cells have an obvious cytoskeleton and bulbous protrusions that suggest vegetative reproduction by budding.[4] Tappania fossils dated near the end of the Boring Billion 0.9-0.8 billion years ago show definite multi-cellularity and differentiation.[4] Some branches that extend from cells appear to re-fuse together, forming net-like patterns.[4] This is a characteristic of fungi, whose hair-like hyphae extending from their cells sometimes fuse in this manner.[4]
First life on land
Plants do not appear on land until 0.47 Ga, but around the end of the Boring Billion (0.7-0.9 Ga) land was beginning to be colonized by cyanobacteria and proto-lichens.[4]
Proto-lichens were symbionts between fungi and algae and left evidence of biological weathering of continental rock in the Doushantuo phosphorite formation (0.6 Ga)[51] and in laterites, Earth's first soils.[52] Additional evidence for life on land is the characteristic carbon isotope signature of photosynthesis that is recorded in rock crusts and ancient soils from the end of the Boring Billion.[53][54] Terrestrial microfossils and microbial mats of cyanobacteria have also been preserved.[55] Some microbial communities lived under rock surfaces, others bound sand together, and some sat atop soils.[56]
See also
References
- 1 2 3 4 5 6 7 Cawood, Peter A.; Hawkesworth, Chris J. (2014-06-01). "Earth's middle age". Geology. 42 (6): 503–506. doi:10.1130/G35402.1. ISSN 0091-7613.
- 1 2 3 Young, Grant M. (2013-05-01). "Precambrian supercontinents, glaciations, atmospheric oxygenation, metazoan evolution and an impact that may have changed the second half of Earth history". Geoscience Frontiers. 4 (3): 247–261. doi:10.1016/j.gsf.2012.07.003.
- ↑ Buick, Roger; Des Marais, David J.; Knoll, Andrew H. (1995-06-20). "Stable isotopic compositions of carbonates from the Mesoproterozoic Bangemall group, northwestern Australia". Chemical Geology. 123 (1–4): 153–171. doi:10.1016/0009-2541(95)00049-R.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 Lenton, Tim; Watson, Andrew. The not-so-boring billion. pp. 242–261. doi:10.1093/acprof:oso/9780199587049.003.0013.
- 1 2 3 4 5 6 Roberts, Nick M. W. (2013-11-01). "The boring billion? – Lid tectonics, continental growth and environmental change associated with the Columbia supercontinent". Geoscience Frontiers. Thematic Section: Antarctica – A window to the far off land. 4 (6): 681–691. doi:10.1016/j.gsf.2013.05.004.
- ↑ Holland, Heinrich D. (2006-06-29). "The oxygenation of the atmosphere and oceans". Philosophical Transactions of the Royal Society of London B: Biological Sciences. 361 (1470): 903–915. doi:10.1098/rstb.2006.1838. ISSN 0962-8436. PMC 1578726
 . PMID 16754606.
. PMID 16754606. - 1 2 3 4 Lyons, Timothy W.; Reinhard, Christopher T.; Planavsky, Noah J. "The rise of oxygen in Earth's early ocean and atmosphere". Nature. 506 (7488): 307–315. doi:10.1038/nature13068.
- ↑ Anbar, A. D.; Knoll, A. H. (2002-08-16). "Proterozoic Ocean Chemistry and Evolution: A Bioinorganic Bridge?". Science. 297 (5584): 1137–1142. doi:10.1126/science.1069651. ISSN 0036-8075. PMID 12183619.
- ↑ Fennel, Katja; Follows, Mick; Falkowski, Paul G. (2005-06-01). "The co-evolution of the nitrogen, carbon and oxygen cycles in the Proterozoic ocean". American Journal of Science. 305 (6-8): 526–545. doi:10.2475/ajs.305.6-8.526. ISSN 0002-9599.
- 1 2 3 Martin, William; Müller, Miklós (1998-03-05). "The hydrogen hypothesis for the first eukaryote". Nature. 392 (6671): 37–41. doi:10.1038/32096. ISSN 0028-0836.
- ↑ Horodyski, R. J.; Knauth, L. P. (1994-01-28). "Life on land in the precambrian". Science (New York, N.Y.). 263 (5146): 494–498. doi:10.1126/science.263.5146.494. ISSN 0036-8075. PMID 17754880.
- ↑ "New fascination with Earth's 'Boring Billion'". Science News. Retrieved 2016-03-01.
- 1 2 Williams, G. E. "Subglacial meltwater channels and glaciofluvial deposits in the Kimberley Basin, Western Australia: 1.8 Ga low-latitude glaciation coeval with continental assembly". Journal of the Geological Society. 162 (1): 111–124. doi:10.1144/0016-764903-157.
- ↑ Brasier, M.D. (1998). "A billion years of environmental stability and the emergence of eukaryotes: New data from northern Australia". Geology.
- ↑ Evans, D. A. D. "Reconstructing pre-Pangean supercontinents". Geological Society of America Bulletin. 125 (11-12): 1735–1751. doi:10.1130/b30950.1.
- ↑ Bradley, Dwight C. (2008-12-01). "Passive margins through earth history". Earth-Science Reviews. 91 (1–4): 1–26. doi:10.1016/j.earscirev.2008.08.001.
- ↑ "When Slime Ruled: Evolutionary Pause Tied to Earth's Stuck Plates". LiveScience.com. Retrieved 2016-02-29.
- ↑ Veizer, Ján (2005-03-01). "Celestial Climate Driver: A Perspective from Four Billion Years of the Carbon Cycle". Geoscience Canada. 32 (1). ISSN 1911-4850.
- ↑ Shaviv, Nir J. (2003-01-01). "The spiral structure of the Milky Way, cosmic rays, and ice age epochs on Earth". New Astronomy. 8 (1): 39–77. doi:10.1016/S1384-1076(02)00193-8.
- 1 2 Planavsky, Noah J.; Reinhard, Christopher T.; Wang, Xiangli; Thomson, Danielle; McGoldrick, Peter; Rainbird, Robert H.; Johnson, Thomas; Fischer, Woodward W.; Lyons, Timothy W. (2014-10-31). "Low Mid-Proterozoic atmospheric oxygen levels and the delayed rise of animals". Science. 346 (6209): 635–638. doi:10.1126/science.1258410. ISSN 0036-8075. PMID 25359975.
- ↑ Eyles, Nick (2008-02-13). "Glacio-epochs and the supercontinent cycle after ∼ 3.0 Ga: Tectonic boundary conditions for glaciation". Palaeogeography, Palaeoclimatology, Palaeoecology. 258 (1–2): 89–129. doi:10.1016/j.palaeo.2007.09.021.
- ↑ Kasting, James F.; Ono, Shuhei (2006-06-29). "Palaeoclimates: the first two billion years". Philosophical Transactions of the Royal Society of London B: Biological Sciences. 361 (1470): 917–929. doi:10.1098/rstb.2006.1839. ISSN 0962-8436. PMC 1868609. PMID 16754607.
- ↑ "Oxygen fluctuations stalled life on Earth". Nature News & Comment. Retrieved 2016-02-24.
- ↑ Partin, C. A.; Bekker, A.; Planavsky, N. J.; Scott, C. T.; Gill, B. C.; Li, C.; Podkovyrov, V.; Maslov, A.; Konhauser, K. O. (2013-05-01). "Large-scale fluctuations in Precambrian atmospheric and oceanic oxygen levels from the record of U in shales". Earth and Planetary Science Letters. 369–370: 284–293. doi:10.1016/j.epsl.2013.03.031.
- ↑ Bekker, A.; Holland, H. D. (2012-02-01). "Oxygen overshoot and recovery during the early Paleoproterozoic". Earth and Planetary Science Letters. 317–318: 295–304. doi:10.1016/j.epsl.2011.12.012.
- ↑ Schröder, S.; Bekker, A.; Beukes, N. J.; Strauss, H.; Van Niekerk, H. S. (2008-04-01). "Rise in seawater sulphate concentration associated with the Paleoproterozoic positive carbon isotope excursion: evidence from sulphate evaporites in the ∼2.2–2.1 Gyr shallow-marine Lucknow Formation, South Africa". Terra Nova. 20 (2): 108–117. doi:10.1111/j.1365-3121.2008.00795.x. ISSN 1365-3121.
- 1 2 "Oxygen fluctuations stalled life on Earth". Nature News & Comment. Retrieved 2016-02-29.
- 1 2 "Evolution stuck in slime for a billion years". phys.org. Retrieved 2016-02-29.
- ↑ Lyons, Timothy W.; Reinhard, Christopher T. (2009-10-27). "An early productive ocean unfit for aerobics". Proceedings of the National Academy of Sciences. 106 (43): 18045–18046. doi:10.1073/pnas.0910345106. ISSN 0027-8424. PMC 2775325. PMID 19846788.
- ↑ Canfield, D. E. (1998-12-03). "A new model for Proterozoic ocean chemistry". Nature. 396 (6710): 450–453. doi:10.1038/24839. ISSN 0028-0836.
- ↑ Boenigk, Jens; Wodniok, Sabina; Glücksman, Edvard (2015-03-31). Biodiversity and Earth History. Springer. ISBN 9783662463949.
- ↑ Johnston, D.T. (2009). "Anoxygenic photosynthesis modulated Proterozoic oxygen and sustained Earth's middle age" (PDF). Proceedings of the National Academy of Sciences.
- ↑ Hecht, Jeff. "Why did evolution stall during the 'boring billion'?". New Scientist. Retrieved 2016-02-23.
- 1 2 "An Overview of Endosymbiotic Models for the Origins of Eukaryotes, Their ATP-Producing Organelles (Mitochondria and Hydrogenosomes), and Their Heterotrophic Lifestyle : Biological Chemistry". www.degruyter.com. Retrieved 2016-02-24.
- 1 2 Mentel, Marek; Martin, William (2008-08-27). "Energy metabolism among eukaryotic anaerobes in light of Proterozoic ocean chemistry". Philosophical Transactions of the Royal Society of London B: Biological Sciences. 363 (1504): 2717–2729. doi:10.1098/rstb.2008.0031. ISSN 0962-8436. PMC 2606767. PMID 18468979.
- 1 2 Searcy, Dennis G.; Hixon, William G. (1991-01-01). "Cytoskeletal origins in sulfur-metabolizing archaebacteria". Biosystems. Papers presented at the 8th Biennial Conference of the International Society for Evolutionary Protistology. 25 (1–2): 1–11. doi:10.1016/0303-2647(91)90008-9.
- ↑ Zang, Wen-Long (2007-06-20). "Deposition and deformation of late Archaean sediments and preservation of microfossils in the Harris Greenstone Domain, Gawler Craton, South Australia". Precambrian Research. 156 (1–2): 107–124. doi:10.1016/j.precamres.2007.03.002.
- ↑ Han, T. M.; Runnegar, B. (1992-07-10). "Megascopic eukaryotic algae from the 2.1-billion-year-old negaunee iron-formation, Michigan". Science. 257 (5067): 232–235. doi:10.1126/science.1631544. ISSN 0036-8075. PMID 1631544.
- ↑ Samuelsson, Joakim; Butterfield, Nicholas J (2001-04-02). "Neoproterozoic fossils from the Franklin Mountains, northwestern Canada: stratigraphic and palaeobiological implications". Precambrian Research. 107 (3–4): 235–251. doi:10.1016/S0301-9268(00)00142-X.
- ↑ Schneider, D. A.; Bickford, M. E.; Cannon, W. F.; Schulz, K. J.; Hamilton, M. A. (2002-06-01). "Age of volcanic rocks and syndepositional iron formations, Marquette Range Supergroup: implications for the tectonic setting of Paleoproterozoic iron formations of the Lake Superior region". Canadian Journal of Earth Sciences. 39 (6): 999–1012. doi:10.1139/e02-016. ISSN 0008-4077.
- ↑ Brocks, Jochen J.; Logan, Graham A.; Buick, Roger; Summons, Roger E. (1999-08-13). "Archean Molecular Fossils and the Early Rise of Eukaryotes". Science. 285 (5430): 1033–1036. doi:10.1126/science.285.5430.1033. ISSN 0036-8075. PMID 10446042.
- ↑ Pearson, Ann (2003). "Phylogenetic and biochemical evidence for sterol synthesis in the bacterium Gemmata obscuriglobus". Proceedings of the National Academy of Sciences. doi:10.1073/pnas.2536559100.
- ↑ Mills, Daniel B.; Canfield, Donald E. (2014-12-01). "Oxygen and animal evolution: Did a rise of atmospheric oxygen "trigger" the origin of animals?". BioEssays. 36 (12): 1145–1155. doi:10.1002/bies.201400101. ISSN 1521-1878.
- ↑ Canfield, Donald (December 3, 1998). "A new model for Proterozoic ocean chemistry". Nature.
- ↑ Reinhard, Christopher T.; Planavsky, Noah J.; Robbins, Leslie J.; Partin, Camille A.; Gill, Benjamin C.; Lalonde, Stefan V.; Bekker, Andrey; Konhauser, Kurt O.; Lyons, Timothy W. (2013-04-02). "Proterozoic ocean redox and biogeochemical stasis". Proceedings of the National Academy of Sciences. 110 (14): 5357–5362. doi:10.1073/pnas.1208622110. ISSN 0027-8424. PMC 3619314. PMID 23515332.
- 1 2 Anbar, A.D. (August 16, 2002). "Proterozoic ocean chemistry and evolution: a bioinorganic bridge". Science.
- ↑ Yan, Y (1993). "Significance of eukaryotic organisms in the microfossil flora of Changeheng system". Acta Micropalaeontologica Sinica.
- ↑ "DNA Repair as the Primary Adaptive Function of Sex in Bacteria and Eukaryotes pp. 1-50". www.novapublishers.com. Retrieved 2016-03-05.
- ↑ Butterfield, Nicholas J. (2000-06-20). "Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes". Paleobiology. 26 (3): 386–404. doi:10.1666/0094-8373(2000)0262.0.CO;2. ISSN 0094-8373.
- ↑ Heckman, Daniel S.; Geiser, David M.; Eidell, Brooke R.; Stauffer, Rebecca L.; Kardos, Natalie L.; Hedges, S. Blair (2001-08-10). "Molecular Evidence for the Early Colonization of Land by Fungi and Plants". Science. 293 (5532): 1129–1133. doi:10.1126/science.1061457. ISSN 0036-8075. PMID 11498589.
- ↑ Anbar, A. D.; Knoll, A. H. (2002-08-16). "Proterozoic Ocean Chemistry and Evolution: A Bioinorganic Bridge?". Science. 297 (5584): 1137–1142. doi:10.1126/science.1069651. ISSN 0036-8075. PMID 12183619.
- ↑ Neaman, Alexander; Chorover, Jon; Brantley, Susan L. (2005-02-01). "Element mobility patterns record organic ligands in soils on early Earth". Geology. 33 (2): 117–120. doi:10.1130/G20687.1. ISSN 0091-7613.
- ↑ Horodyski, R. J.; Knauth, L. P. (1994-01-28). "Life on land in the precambrian". Science (New York, N.Y.). 263 (5146): 494–498. doi:10.1126/science.263.5146.494. ISSN 0036-8075. PMID 17754880.
- ↑ Retallack, Gregory J.; Mindszenty, Andrea (1994-04-01). "Well preserved late Precambrian Paleosols from Northwest Scotland". Journal of Sedimentary Research. 64 (2a): 264–281. doi:10.1306/D4267D7A-2B26-11D7-8648000102C1865D. ISSN 1527-1404.
- ↑ Watanabe, Yumiko; Martini, Jacques E. J.; Ohmoto, Hiroshi (2000-11-30). "Geochemical evidence for terrestrial ecosystems 2.6 billion years ago". Nature. 408 (6812): 574–578. doi:10.1038/35046052. ISSN 0028-0836.
- ↑ "Life on land in the Proterozoic: Evidence from the Torridonian rocks of northwest Scotland - University of St Andrews". risweb.st-andrews.ac.uk. Retrieved 2016-03-05.