Luciferase
| Firefly luciferase | |
|---|---|
|
Structure of Photinus pyralis firefly luciferase. | |
| Identifiers | |
| Organism | |
| Symbol | Firefly luciferase |
| PDB | 1LCI More structures |
| UniProt | P08659 |
| Other data | |
| EC number | 1.13.12.7 |
| Bacterial Luciferase monooxygenase family | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | Bac_luciferase | ||||||||
| Pfam | PF00296 | ||||||||
| InterPro | IPR016048 | ||||||||
| PROSITE | PDOC00397 | ||||||||
| SCOP | 1nfp | ||||||||
| SUPERFAMILY | 1nfp | ||||||||
| |||||||||
| Dinoflagellate Luciferase catalytic domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 crystal structure of a luciferase domain from the dinoflagellate Lingulodinium polyedrum | |||||||||
| Identifiers | |||||||||
| Symbol | Luciferase_cat | ||||||||
| Pfam | PF10285 | ||||||||
| InterPro | IPR018804 | ||||||||
| |||||||||
| Dinoflagellate Luciferase/LBP N-terminal domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | Luciferase_N | ||||||||
| Pfam | PF05295 | ||||||||
| InterPro | IPR007959 | ||||||||
| |||||||||
| Dinoflagellate Luciferase helical bundle domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
|
crystal structure of a Dinoflagellate luciferase domain from the dinoflagellate Lingulodinium polyedrum | |||||||||
| Identifiers | |||||||||
| Symbol | Luciferase_3H | ||||||||
| Pfam | PF10284 | ||||||||
| InterPro | IPR018475 | ||||||||
| |||||||||
Luciferase is a generic term for the class of oxidative enzymes that produce bioluminescence, and is distinct from a photoprotein. The name is derived from Lucifer, the root of which means 'light-bearer' (lucem ferre). One example is the firefly luciferase (EC 1.13.12.7) from the firefly Photinus pyralis.[1] "Firefly luciferase" as a laboratory reagent often refers to P. pyralis luciferase although recombinant luciferases from several other species of fireflies are also commercially available.
Examples
A variety of organisms regulate their light production using different luciferases in a variety of light-emitting reactions. The most famous are the fireflies,[2] although the enzyme exists in organisms as different as the Jack-O-Lantern mushroom (Omphalotus olearius) and many marine creatures.
Firefly and click beetle
The luciferases of fireflies - of which there are over 2000 species - and of the other Elateroidea (click beetles and relatives in general) are diverse enough to be useful in molecular phylogeny. In fireflies, the oxygen required is supplied through a tube in the abdomen called the abdominal trachea. One well-studied luciferase is that of the Photinini firefly Photinus pyralis, which has an optimum pH of 7.8.[3]
Sea pansy
Also well studied is the fancy sea pansy, Renilla reniformis. In this organism, the luciferase (Renilla-luciferin 2-monooxygenase) is closely associated with a luciferin-binding protein as well as a green fluorescent protein (GFP). Calcium triggers release of the luciferin (coelenterazine) from the luciferin binding protein. The substrate is then available for oxidation by the luciferase, where it is degraded to coelenteramide with a resultant release of energy. In the absence of GFP, this energy would be released as a photon of blue light (peak emission wavelength 482 nm). However, due to the closely associated GFP, the energy released by the luciferase is instead coupled through resonance energy transfer to the fluorophore of the GFP, and is subsequently released as a photon of green light (peak emission wavelength 510 nm). The catalyzed reaction is:[4]
- coelenterazine + O2 → coelenteramide + CO2 + photon of light
Bacterial
Bacterial bioluminescence is seen in Photobacterium species, Vibrio fischeri, Vibrio haweyi, and Vibrio harveyi. Light emission in some bioluminescent bacteria utilizes 'antenna' such as 'lumazine protein' to accept the energy from the primary excited state on the luciferase, resulting in an excited lulnazine chromophore which emits light that is of a shorter wavelength (more blue), while in others use a yellow fluorescent protein (YFP) with FMN as the chromophore and emits light that is red-shifted relative to that from luciferase.[5]
Dinoflagellate
Dinoflagellate luciferase is a multi-domain protein, consisting of an N-terminal domain, and three catalytic domains, each of which preceded by a helical bundle domain. The structure of the dinoflagellate luciferase catalytic domain has been solved.[6] The core part of the domain is a 10 stranded beta barrel that is structurally similar to lipocalins and FABP.[6] The N-terminal domain is conserved between dinoflagellate luciferase and luciferin binding proteins (LBPs). It has been suggested that this region may mediate an interaction between LBP and luciferase or their association with the vacuolar membrane.[7] The helical bundle domain has a three helix bundle structure that holds four important histidines that are thought to play a role in the pH regulation of the enzyme.[6] There is a large pocket in the β-barrel of the dinoflagellate luciferase at pH 8 to accommodate the tetrapyrrole substrate but there is no opening to allow the substrate to enter. Therefore, a significant conformational change must occur to provide access and space for a ligand in the active site and the source for this change is through the four N-terminal histidine residues.[6] At pH 8, it can be seen that the unprotonated histidine residues are involved in a network of hydrogen bonds at the interface of the helices in the bundle that block substrate access to the active site and disruption of this interaction by protonation (at pH 6.3) or by replacement of the histidine residues by alanine causes a large molecular motion of the bundle, separating the helices by 11Å and opening the catalytic site.[6] Logically, the histidine residues cannot be replaced by alanine in nature but this experimental replacement further confirms that the larger histidine residues block the active site. Additionally, three Gly-Gly sequences, one in the N-terminal helix and two in the helix-loop-helix motif, could serve as hinges about which the chains rotate in order to further open the pathway to the catalytic site and enlarge the active site.[6]
A dinoflagellate luciferase is capable of emitting light due to its interaction with its substrate (luciferin) and the luciferin-binding protein (LBP) in the scintillon organelle found in dinoflagellates.[6] The luciferase acts in accordance with luciferin and LBP in order to emit light but each component functions at a different pH. Luciferase and its domains are not active at pH 8 but they are extremely active at the optimum pH of 6.3 whereas LBP binds luciferin at pH 8 and releases it at pH 6.3.[6] Consequently, luciferin is only released to react with an active luciferase when the scintillon is acidified to pH 6.3. Therefore, in order to lower the pH, voltage-gated channels in the scintillon membrane are opened to allow the entry of protons from a vacuole possessing an action potential produced from a mechanical stimulation.[6] Hence, it can be seen that the action potential in the vacuolar membrane leads to acidification and this in turn allows the luciferin to be released to react with luciferase in the scintillon, producing a flash of blue light.
Copepod
Newer luciferases have recently been identified that, unlike other luciferases above, are naturally secreted molecules. One such example is the Metridia luciferase (MetLuc) that is derived from the marine copepod Metridia longa. The Metridia longa secreted luciferase gene encodes a 24 kDa protein containing an N-terminal secretory signal peptide of 17 amino acid residues. The sensitivity and high signal intensity of this luciferase molecule proves advantageous in many reporter studies. Some of the benefits of using a secreted reporter molecule like MetLuc is its no-lysis protocol that allows one to be able to conduct live cell assays and multiple assays on the same cell.[8]
Mechanism of reaction
The chemical reaction catalyzed by firefly luciferase takes place in two steps:
- luciferin + ATP → luciferyl adenylate + PPi
- luciferyl adenylate + O2 → oxyluciferin + AMP + light
Light is produced because the reaction forms oxyluciferin in an electronically excited state. The reaction releases a photon of light as oxyluciferin goes back to the ground state.
Luciferyl adenylate can additionally participate in a side reaction with O2 to form hydrogen peroxide and dehydroluciferyl-AMP. About 20% of the luciferyl adenylate intermediate is oxidized in this pathway.[9]
The reaction catalyzed by bacterial luciferase is also an oxidative process:
- FMNH2 + O2 + RCHO → FMN + RCOOH + H2O + light
In the reaction, a reduced flavin mononucleotide oxidizes a long-chain aliphatic aldehyde to an aliphatic carboxylic acid. The reaction forms an excited hydroxyflavin intermediate, which is dehydrated to the product FMN to emit blue-green light.[10]
Nearly all of the energy input into the reaction is transformed into light. The reaction is 80%[11] to 90%[12] efficient. As a comparison, the incandescent light bulb only converts about 10% of its energy into light.[13] and a 150 lumen per Watt (lm/W) LED converts 20% of input energy to visible light.[12]
Firefly luciferase generates light from luciferin in a multistep process. First, D-luciferin is adenylated by MgATP to form luciferyl adenylate and pyrophosphate. After activation by ATP, luciferyl adenylate is oxidized by molecular oxygen to form a dioxetanone ring. A decarboxylation reaction forms an excited state of oxyluciferin, which tautomerizes between the keto-enol form. The reaction finally emits light as oxyluciferin returns to the ground state.[2]


Bifunctionality
Luciferase can function in two different pathways: a bioluminescence pathway and a CoA-ligase pathway.[15] In both pathways, luciferase initially catalyzes an adenylation reaction with MgATP. However, in the CoA-ligase pathway, CoA can displace AMP to form luciferyl CoA.
Fatty acyl-CoA synthetase similarly activates fatty acids with ATP, followed by displacement of AMP with CoA. Because of their similar activities, luciferase is able to replace fatty acyl-CoA synthetase and convert long-chain fatty acids into fatty-acyl CoA for beta oxidation.[15]
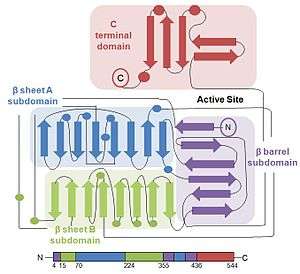
Structure
The protein structure of firefly luciferase consists of two compact domains: the N-terminal domain and the C-terminal domain. The N-terminal domain is composed of two β-sheets in an αβαβα structure and a β barrel. The two β-sheets stack on top of each other, with the β-barrel covering the end of the sheets.[2]
The C-terminal domain is connected to the N-terminal domain by a flexible hinge, which can separate the two domains. The amino acid sequences on the surface of the two domains facing each other are conserved in bacterial and firefly luciferase, thereby strongly suggesting that the active site is located in the cleft between the domains.[16]
During a reaction, luciferase has a conformational change and goes into a “closed” form with the two domains coming together to enclose the substrate. This ensures that water is excluded from the reaction and does not hydrolyze ATP or the electronically excited product.[16]
Spectral differences in bioluminescence
Firefly luciferase bioluminescence color can vary between yellow-green (λmax = 550 nm) to red (λmax = 620).[17] There are currently several different mechanisms describing how the structure of luciferase affects the emission spectrum of the photon and effectively the color of light emitted.
One mechanism proposes that the color of the emitted light depends on whether the product is in the keto or enol form. The mechanism suggests that red light is emitted from the keto form of oxyluciferin, while green light is emitted from the enol form of oxyluciferin.[18][19] However, 5,5-dimethyloxyluciferin emits green light even though it is constricted to the keto form because it cannot tautomerize.[20]
Another mechanism proposes that twisting the angle between benzothiazole and thiazole rings in oxyluciferin determines the color of bioluminescence. This explanation proposes that a planar form with an angle of 0° between the two rings corresponds to a higher energy state and emits a higher-energy green light, whereas an angle of 90° puts the structure in a lower energy state and emits a lower-energy red light.[21]
The most recent explanation for the bioluminescence color examines the microenvironment of the excited oxyluciferin. Studies suggest that the interactions between the excited state product and nearby residues can force the oxyluciferin into an even higher energy form, which results in the emission of green light. For example, Arg 218 has electrostatic interactions with other nearby residues, restricting oxyluciferin from tautomerizing to the enol form.[22] Similarly, other results have indicated that the microenvironment of luciferase can force oxyluciferin into a more rigid, high-energy structure, forcing it to emit a high-energy green light.[23]
Regulation
D-luciferin is the substrate for firefly luciferase’s bioluminescence reaction, while L-luciferin is the substrate for luciferyl-CoA synthetase activity. Both reactions are inhibited by the substrate’s enantiomer: L-luciferin and D-luciferin inhibit the bioluminescence pathway and the CoA-ligase pathway, respectively.[14] This shows that luciferase can differentiate between the isomers of the luciferin structure.
L-luciferin is able to emit a weak light even though it is a competitive inhibitor of D-luciferin and the bioluminescence pathway.[24] Light is emitted because the CoA synthesis pathway can be converted to the bioluminescence reaction by hydrolyzing the final product via an esterase back to D-luciferin.[14]
Luciferase activity is additionally inhibited by oxyluciferin[25] and allosterically activated by ATP. When ATP binds to the enzyme’s two allosteric sites, luciferase’s affinity to bind ATP in its active site increases.[17]
Applications
Luciferase can be produced in the lab through genetic engineering for a number of purposes. Luciferase genes can be synthesized and inserted into organisms or transfected into cells. Mice, silkworms, and potatoes are just a few of the organisms that have already been engineered to produce the protein.[26]
In the luciferase reaction, light is emitted when luciferase acts on the appropriate luciferin substrate. Photon emission can be detected by light sensitive apparatus such as a luminometer or modified optical microscopes. This allows observation of biological processes.[27] Since light excitation is not needed for luciferase bioluminescence, there is minimal autofluorescence and therefore virtually background-free fluorescence.[28] Therefore, as little as 0.02pg can still be accurately measured using a standard scintillation counter.[29]
In biological research, luciferase is commonly used as a reporter to assess the transcriptional activity in cells that are transfected with a genetic construct containing the luciferase gene under the control of a promoter of interest.[30][31][32] Additionally proluminescent molecules that are converted to luciferin upon activity of a particular enzyme can be used to detect enzyme activity in coupled or two-step luciferase assays. Such substrates have been used to detect caspase activity and cytochrome P450 activity, among others.[27][30]
Luciferase can also be used to detect the level of cellular ATP in cell viability assays or for kinase activity assays.[30][33] Luciferase can act as an ATP sensor protein through biotinylation. Biotinylation will immobilize luciferase on the cell-surface by binding to a streptavidin-biotin complex. This allows luciferase to detect the efflux of ATP from the cell and will effectively display the real-time release of ATP through bioluminescence.[34] Luciferase can additionally be made more sensitive for ATP detection by increasing the luminescence intensity by changing certain amino acid residues in the sequence of the protein.[35]
Whole animal imaging (referred to as in vivo or, occasionally, ex vivo imaging) is a powerful technique for studying cell populations in live animals, such as mice.[36] Different types of cells (e.g. bone marrow stem cells, T-cells) can be engineered to express a luciferase allowing their non-invasive visualization inside a live animal using a sensitive charge-couple device camera (CCD camera).This technique has been used to follow tumorigenesis and response of tumors to treatment in animal models.[37][38] However, environmental factors and therapeutic interferences may cause some discrepancies between tumor burden and bioluminescence intensity in relation to changes in proliferative activity. The intensity of the signal measured by in vivo imaging may depend on various factors, such as D-luciferin absorption through the peritoneum, blood flow, cell membrane permeability, availability of co-factors, intracellular pH and transparency of overlying tissue, in addition to the amount of luciferase.[39]
The Glowing Plant project plans to use bacterial bio-luminescent systems to engineer novelty glowing Arabidopsis thaliana plants. Longer term they hypothesize that maybe such systems could be used to create eco-friendly sustainable light sources.[40]
Luciferase is a heat-sensitive protein that is used in studies on protein denaturation, testing the protective capacities of heat shock proteins. The opportunities for using luciferase continue to expand.[41]
See also
- Firefly
- Luciferin
- Firefly luciferin
- Bioluminescence imaging
- Quorum sensing
- Bioluminescent bacteria
- Mouse models of breast cancer metastasis
References
- ↑ Gould SJ, Subramani S (Nov 1988). "Firefly luciferase as a tool in molecular and cell biology". Analytical Biochemistry. 175 (1): 5–13. doi:10.1016/0003-2697(88)90353-3. PMID 3072883.
- 1 2 3 4 Baldwin TO (Mar 1996). "Firefly luciferase: the structure is known, but the mystery remains". Structure. 4 (3): 223–8. doi:10.1016/S0969-2126(96)00026-3. PMID 8805542.
- ↑ Steghens JP, Min KL, Bernengo JC (Nov 1998). "Firefly luciferase has two nucleotide binding sites: effect of nucleoside monophosphate and CoA on the light-emission spectra". The Biochemical Journal. 336 ( Pt 1) (1): 109–13. PMC 1219848
 . PMID 9806891.
. PMID 9806891. - ↑ Shimomura O (1985). "Bioluminescence in the sea: photoprotein systems". Symposia of the Society for Experimental Biology. 39: 351–72. PMID 2871634.
- ↑ Baldwin TO, Christopher JA, Raushel FM, Sinclair JF, Ziegler MM, Fisher AJ, Rayment I (Dec 1995). "Structure of bacterial luciferase". Current Opinion in Structural Biology. 5 (6): 798–809. doi:10.1016/0959-440x(95)80014-x. PMID 8749369.
- 1 2 3 4 5 6 7 8 9 Schultz LW, Liu L, Cegielski M, Hastings JW (Feb 2005). "Crystal structure of a pH-regulated luciferase catalyzing the bioluminescent oxidation of an open tetrapyrrole". Proceedings of the National Academy of Sciences of the United States of America. 102 (5): 1378–83. doi:10.1073/pnas.0409335102. PMC 547824. PMID 15665092.
- ↑ Okamoto OK, Liu L, Robertson DL, Hastings JW (Dec 2001). "Members of a dinoflagellate luciferase gene family differ in synonymous substitution rates". Biochemistry. 40 (51): 15862–8. doi:10.1021/bi011651q. PMID 11747464.
- ↑ Huh S, Lee J, Jung E, Kim SC, Kang JI, Lee J, Kim YW, Sung YK, Kang HK, Park D (Jun 2009). "A cell-based system for screening hair growth-promoting agents". Archives of Dermatological Research. 301 (5): 381–5. doi:10.1007/s00403-009-0931-0. PMID 19277688.
- ↑ Fraga H, Fernandes D, Novotny J, Fontes R, Esteves da Silva JC (Jun 2006). "Firefly luciferase produces hydrogen peroxide as a coproduct in dehydroluciferyl adenylate formation". Chembiochem. 7 (6): 929–35. doi:10.1002/cbic.200500443. PMID 16642538.
- ↑ Fisher AJ, Thompson TB, Thoden JB, Baldwin TO, Rayment I (Sep 1996). "The 1.5-A resolution crystal structure of bacterial luciferase in low salt conditions". The Journal of Biological Chemistry. 271 (36): 21956–68. doi:10.1074/jbc.271.36.21956. PMID 8703001.
- ↑ Elizabeth Wilson (Jan 18, 1999). "What's That Stuff?". Chemical and Engineering News. 77 (3): 65. doi:10.1021/cen-v077n003.p065.
- 1 2 Vanessa Knivett (2009). "Lighting the way". EE times.
- ↑ General Electric TP-110, page 23, table.
- 1 2 3 Nakamura M, Maki S, Amano Y, Ohkita Y, Niwa K, Hirano T, Ohmiya Y, Niwa H (Jun 2005). "Firefly luciferase exhibits bimodal action depending on the luciferin chirality". Biochemical and Biophysical Research Communications. 331 (2): 471–5. doi:10.1016/j.bbrc.2005.03.202. PMID 15850783.
- 1 2 Oba Y, Ojika M, Inouye S (Apr 2003). "Firefly luciferase is a bifunctional enzyme: ATP-dependent monooxygenase and a long chain fatty acyl-CoA synthetase". FEBS Letters. 540 (1-3): 251–4. doi:10.1016/S0014-5793(03)00272-2. PMID 12681517.
- 1 2 3 Conti E, Franks NP, Brick P (Mar 1996). "Crystal structure of firefly luciferase throws light on a superfamily of adenylate-forming enzymes". Structure. 4 (3): 287–98. doi:10.1016/S0969-2126(96)00033-0. PMID 8805533.
- 1 2 Ugarova NN (Jul 1989). "Luciferase of Luciola mingrelica fireflies. Kinetics and regulation mechanism". Journal of Bioluminescence and Chemiluminescence. 4 (1): 406–18. doi:10.1002/bio.1170040155. PMID 2801227.
- ↑ White EH, Rapaport E, Hopkins TA, Seliger HH (Apr 1969). "Chemi- and bioluminescence of firefly luciferin". Journal of the American Chemical Society. 91 (8): 2178–80. doi:10.1021/ja01036a093. PMID 5784183.
- ↑ Fraga H (Feb 2008). "Firefly luminescence: a historical perspective and recent developments". Photochemical & Photobiological Sciences. 7 (2): 146–58. doi:10.1039/b719181b. PMID 18264582.
- ↑ Branchini BR, Southworth TL, Murtiashaw MH, Magyar RA, Gonzalez SA, Ruggiero MC, Stroh JG (Jun 2004). "An alternative mechanism of bioluminescence color determination in firefly luciferase". Biochemistry. 43 (23): 7255–62. doi:10.1021/bi036175d. PMID 15182171.
- ↑ McCapra F, Gilfoyle DJ, Young DW, et al. (1994). Bioluminescence and Chemiluminescence: Fundamentals and Applied.
- ↑ Nakatani N, Hasegawa JY, Nakatsuji H (Jul 2007). "Red light in chemiluminescence and yellow-green light in bioluminescence: color-tuning mechanism of firefly, Photinus pyralis, studied by the symmetry-adapted cluster-configuration interaction method". Journal of the American Chemical Society. 129 (28): 8756–65. doi:10.1021/ja0611691. PMID 17585760.
- ↑ Nakamura M, Niwa K, Maki S, et al. (December 2006). "Construction of a new firefly bioluminescence system using L-luciferin as substrate". Anal. Biochem. 47 (1): 1197–1200. doi:10.1016/j.tetlet.2005.12.033.
- ↑ Lembert N (Jul 1996). "Firefly luciferase can use L-luciferin to produce light". The Biochemical Journal. 317 ( Pt 1) (1): 273–7. PMC 1217473. PMID 8694774.
- ↑ Ribeiro C, Esteves da Silva JC (Sep 2008). "Kinetics of inhibition of firefly luciferase by oxyluciferin and dehydroluciferyl-adenylate". Photochemical & Photobiological Sciences. 7 (9): 1085–90. doi:10.1039/b809935a. PMID 18754056.
- ↑ Contag CH, Bachmann MH (2002). "Advances in in vivo bioluminescence imaging of gene expression". Annual Review of Biomedical Engineering. 4: 235–60. doi:10.1146/annurev.bioeng.4.111901.093336. PMID 12117758.
- 1 2 "Introduction to Bioluminescence Assays". Promega Corporation. Retrieved 2009-03-07.
- ↑ Williams TM, Burlein JE, Ogden S, Kricka LJ, Kant JA (Jan 1989). "Advantages of firefly luciferase as a reporter gene: application to the interleukin-2 gene promoter". Analytical Biochemistry. 176 (1): 28–32. doi:10.1016/0003-2697(89)90267-4. PMID 2785354.
- ↑ Nguyen VT, Morange M, Bensaude O (Jun 1988). "Firefly luciferase luminescence assays using scintillation counters for quantitation in transfected mammalian cells". Analytical Biochemistry. 171 (2): 404–8. doi:10.1016/0003-2697(88)90505-2. PMID 3407940.
- 1 2 3 Fan F, Wood KV (Feb 2007). "Bioluminescent assays for high-throughput screening". ASSAY and Drug Development Technologies. 5 (1): 127–36. doi:10.1089/adt.2006.053. PMID 17355205.
- ↑ Rozema E, Atanasov AG, Fakhrudin N, Singhuber J, Namduang U, Heiss EH, Reznicek G, Huck CW, Bonn GK, Dirsch VM, Kopp B (2012). "Selected Extracts of Chinese Herbal Medicines: Their Effect on NF-κB, PPARα and PPARγ and the Respective Bioactive Compounds". Evidence-Based Complementary and Alternative Medicine. 2012: 983023. doi:10.1155/2012/983023. PMC 3366346. PMID 22675394.
- ↑ http://altogenlabs.com/services/generation_of_stable_cell_lines/stable_cells_generation
- ↑ Meisenheimer PL, O'Brien MA, Cali JJ (September 2008). "Luminogenic enzyme substrates: The basis for a new paradigm in assay design." (PDF). Promega Notes. 100: 22–26.
- ↑ Nakamura M, Mie M, Funabashi H, Yamamoto K, Ando J, Kobatake E (May 2006). "Cell-surface-localized ATP detection with immobilized firefly luciferase". Analytical Biochemistry. 352 (1): 61–7. doi:10.1016/j.ab.2006.02.019. PMID 16564487.
- ↑ Fujii H, Noda K, Asami Y, Kuroda A, Sakata M, Tokida A (Jul 2007). "Increase in bioluminescence intensity of firefly luciferase using genetic modification". Analytical Biochemistry. 366 (2): 131–6. doi:10.1016/j.ab.2007.04.018. PMID 17540326.
- ↑ Greer LF, Szalay AA (2002). "Imaging of light emission from the expression of luciferases in living cells and organisms: a review". Luminescence. 17 (1): 43–74. doi:10.1002/bio.676. PMID 11816060.
- ↑ Lyons SK, Meuwissen R, Krimpenfort P, Berns A (Nov 2003). "The generation of a conditional reporter that enables bioluminescence imaging of Cre/loxP-dependent tumorigenesis in mice". Cancer Research. 63 (21): 7042–6. PMID 14612492.
- ↑ Becher OJ, Holland EC (Apr 2006). "Genetically engineered models have advantages over xenografts for preclinical studies". Cancer Research. 66 (7): 3355–8, discussion 3358–9. doi:10.1158/0008-5472.CAN-05-3827. PMID 16585152.
- ↑ Inoue Y, Tojo A, Sekine R, Soda Y, Kobayashi S, Nomura A, Izawa K, Kitamura T, Okubo T, Ohtomo K (May 2006). "In vitro validation of bioluminescent monitoring of disease progression and therapeutic response in leukaemia model animals". European Journal of Nuclear Medicine and Molecular Imaging. 33 (5): 557–65. doi:10.1007/s00259-005-0048-4. PMID 16501974.
- ↑ http://www.kickstarter.com/projects/antonyevans/glowing-plants-natural-lighting-with-no-electricit
- ↑ Massoud TF, Paulmurugan R, De A, Ray P, Gambhir SS (Feb 2007). "Reporter gene imaging of protein-protein interactions in living subjects". Current Opinion in Biotechnology. 18 (1): 31–7. doi:10.1016/j.copbio.2007.01.007. PMID 17254764.
External links
-
 Media related to Luciferase at Wikimedia Commons
Media related to Luciferase at Wikimedia Commons - Trimmer B, Zayas R, Qazi S, Lewis S, Michel T, Dudzinski D, Aprille J, Lagace C (2001-06-28). "Firefly flashes and Nitric Oxide". Tufts University. Retrieved 2008-10-02.
- "Trends in development of reporter genes". reportergene.com. Retrieved 2009-03-07.
- "BL Web: Luciferin types". The Bioluminescence Web Page. University of California, Santa Barbara. Retrieved 2009-03-07.
- "Bioluminescence Reporters Protocols and Applications Guide". Protocols and applications. Promega Corporation. Retrieved 2009-03-07.
- "BL Web: Luciferin types". ISCID Encyclopedia of Science and Philosophy. ISCID. Retrieved 2010-04-20.
- David Goodsell. "Luciferase". Molecule of the Month. Protein Data Bank. Retrieved 2013-01-15.
This article incorporates text from the public domain Pfam and InterPro IPR018804 This article incorporates text from the public domain Pfam and InterPro IPR007959 This article incorporates text from the public domain Pfam and InterPro IPR018475