Founder effect
In population genetics, the founder effect is the loss of genetic variation that occurs when a new population is established by a very small number of individuals from a larger population. It was first fully outlined by Ernst Mayr in 1942,[1] using existing theoretical work by those such as Sewall Wright.[2] As a result of the loss of genetic variation, the new population may be distinctively different, both genotypically and phenotypically, from the parent population from which it is derived. In extreme cases, the founder effect is thought to lead to the speciation and subsequent evolution of new species.
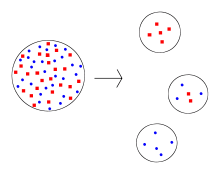
In the figure shown, the original population has nearly equal numbers of blue and red individuals. The three smaller founder populations show that one or the other color may predominate (founder effect), due to random sampling of the original population. A population bottleneck may also cause a founder effect, though it is not strictly a new population.
The founder effect occurs when a small group of migrants that is not genetically representative of the population from which they came establish in a new area.[3][4] In addition to founder effects, the new population is often a very small population, so shows increased sensitivity to genetic drift, an increase in inbreeding, and relatively low genetic variation. This can be observed in the limited gene pools of Icelanders, Ashkenazi Jews, Faroe Islanders, Easter Islanders, and those native to Pitcairn Island. Another example is the remarkably high deaf population of Martha's Vineyard, which resulted in the development of Martha's Vineyard Sign Language.
Founder mutation
In genetics, a founder mutation is a mutation that appears in the DNA of one or more individuals which are founders of a distinct population. Founder mutations initiate with changes that occur in the DNA and can be passed down to other generations.[5][6]
Founder mutations originate in long stretches of DNA on a single chromosome; indeed, the original haplotype is the whole chromosome. As the generations progress, the proportion of the haplotype that is common to all carriers of the mutation is shortened (due to genetic recombination). This shortening allows scientists to roughly estimate the age of the mutation.[7]
General
The founder effect is a special case of genetic drift, occurring when a small group in a population splinters off from the original population and forms a new one. The new colony may have less genetic variation than the original population, and through the random sampling of alleles during reproduction of subsequent generations, continue rapidly towards fixation. This consequence of inbreeding makes the colony more vulnerable to extinction.
When a newly formed colony is small, its founders can strongly affect the population's genetic makeup far into the future. In humans, which have a slow reproduction rate, the population will remain small for many generations, effectively amplifying the drift effect generation after generation until the population reaches a certain size. Alleles which were present but relatively rare in the original population can move to one of two extremes. The most common one is that the allele is soon lost altogether, but the other possibility is that the allele survives and within a few generations has become much more dispersed throughout the population. The new colony can experience an increase in the frequency of recessive alleles, as well, and as a result, an increased number who are homozygous for certain recessive traits.
The variation in gene frequency between the original population and colony may also trigger the two groups to diverge significantly over the course of many generations. As the variance, or genetic distance, increases, the two separated populations may become distinctively different, both genetically and phenotypically, although not only genetic drift, but also natural selection, gene flow and mutation all contribute to this divergence. This potential for relatively rapid changes in the colony's gene frequency led most scientists to consider the founder effect (and by extension, genetic drift) a significant driving force in the evolution of new species. Sewall Wright was the first to attach this significance to random drift and small, newly isolated populations with his shifting balance theory of speciation.[8] Following behind Wright, Ernst Mayr created many persuasive models to show that the decline in genetic variation and small population size accompanying the founder effect were critically important for new species to develop.[9] However, much less support for this view is shown today, since the hypothesis has been tested repeatedly through experimental research, and the results have been equivocal at best. Speciation by genetic drift is a specific case of peripatric speciation which in itself occurs in rare instances.[10] It takes place when a random change in genetic frequency of population favours the survival of a few organisms of the species with rare genes which cause reproductive mutation. These surviving organisms then breed among themselves over a long period of time to create a whole new species whose reproductive systems or behaviors are no more compatible with the original population. [11]
Serial founder effect
Serial founder effects have occurred when populations migrate over long distances. Such long-distance migrations typically involve relatively rapid movements followed by periods of settlement. The populations in each migration carry only a subset of the genetic diversity carried from previous migrations. As a result, genetic differentiation tends to increase with geographic distance as described by the "isolation by distance" model.[12] The migration of humans out of Africa is characterized by serial founder effects.[13] Africa has the highest degree of genetic diversity of any continent, which is consistent with an African origin of modern humans. After the initial migration from Africa, the Indian subcontinent was the first major settling point for modern humans. Consequently, India has the second-highest genetic diversity in the world. In general, the genetic diversity of the Indian subcontinent is a subset of Africa, and the genetic diversity outside Africa is a subset of India.[14][15]
Founder effects in island ecology
Founder populations are essential to the study of island biogeography and island ecology. A natural "blank slate" is not easily found, but a classic series of studies on founder population effects was done following the catastrophic 1883 eruption of Krakatoa, which erased all life on the island. Another continuing study has been following the biocolonization of Surtsey, Iceland, a new volcanic island that erupted offshore between 1963 and 1967. An earlier event, the Toba eruption in Sumatra about 73,000 years ago, covered some parts of India with 3–6 m (10–20 ft) of ash, and must have coated the Nicobar Islands and Andaman Islands, much nearer in the ash fallout cone, with life-smothering layers, forcing the restart of their biodiversity.
However, not all founder effect studies are initiated after a natural disaster; some scientists study the reinstatement of a species that became locally extinct. Hajji and others[16] and Hundertmark & Van Daele [17] studied the current population statuses of past founder effects in Corsican red deer and Alaskan elk, respectively. Corsican red deer are still listed as an endangered species, decades after a severe bottleneck. They inhabit the Tryrrhenian islands and surrounding mainlands currently, and before the bottleneck, but Hajji and others wanted to know how the deer originally got to the islands, and from what parent population or species they were derived. Through molecular analysis, they were able to determine a possible lineage, with red deer from the islands of Corsica and Sardinia being the most related to one another. These results are promising, as the island of Corsica was repopulated with red deer from the Sardinian island after the original Corsican red deer population became extinct, and the deer now inhabiting the island of Corsica are diverging from those inhabiting Sardinia. Its founder effect consequences are still a work in progress.
Hundertmark and Van Daele examined the genetic differences between a parent population of elk of Washington on the continental USA, and the growing population fertilized by eight founding elk on an Alaskan island. After 80 years, this island population has genetically and allelically diverged significantly from the parent population in Washington, as a result of the founding individuals’ lesser genetic diversity. When statistically analyzing the rapid growth of the island population, Hundertmark and Van Daele came to the conclusion, “…that a severe bottleneck followed by rapid population growth may be undetectable using available tests.” Kolbe and others [18] set up a pair of genetically sequenced and morphologically examined lizards on seven small islands to watch each new population’s growth and adaptation to its new environment. Specifically, they were looking at the effects on limb length and perch width, both widely varying phenotypic ranges in the parent population. Unfortunately, immigration did occur, but the founder effect and adaptive differentiation, which could eventually lead to peripatric speciation, were statistically and biologically significant between the island populations after a few years. The authors also point out that although adaptive differentiation is significant, the differences between island populations best reflect the differences between founders and their genetic diversity that has been passed down through the generations.
Founder effects in human populations
Due to various migrations throughout human history, founder effects are somewhat common among humans in different times and places. The French Canadians of Quebec are a classical example of founder population. Over 150 years of French colonization, between 1608 and 1760, an estimated 8,500 pioneers married and left at least one descendant on the territory.[19] Following the takeover of the colony by the British crown in 1760, immigration from France effectively stopped, but descendents of French settlers continued to grow in number mainly because of high fertility rate. Intermarriage occurred mostly with the deported Acadians and migrants coming from the British Isles. Since the 20th century, immigration in Quebec and mixing of French Canadians involve people from all over the world. While the French Canadians of Quebec today may be partly of other ancestries, the genetic contribution of the original French founders is predominant, explaining about 90% of regional gene pools, while Acadians (descended from other French settlers in eastern Canada) explain 4%, British 2% and Native American and other groups contributed less.[20]
Founder effects can also occur naturally as competing genetic lines die out. This means that an effective founder population consists only of those whose genetic print is identifiable in subsequent populations. Because in sexual reproduction, genetic recombination ensures that with each generation, only half the genetic material of a parent is represented in the offspring, some genetic lines may die out entirely, though numerous progeny are born. The misinterpretations of "Mitochondrial Eve" are a case in point: it may be hard to explain that a "mitochondrial Eve" was not the only woman of her time.
In humans, founder effects can arise from cultural isolation, and inevitably, endogamy. For example, the Amish populations in the United States exhibit founder effects because they have grown from a very few founders, have not recruited newcomers, and tend to marry within the community. Though still rare, phenomena such as polydactyly (extra fingers and toes, a symptom of Ellis-van Creveld syndrome) are more common in Amish communities than in the American population at large.[21] Maple syrup urine disease affects about one out of 180,000 infants in the general population.[22] Due in part to the founder effect,[23] however, the disease has a much higher prevalence in children of Amish, Mennonite, and Jewish descent.[24][22][25] Similarly, a high frequency of fumarase deficiency exists among the 10,000 members of the Fundamentalist Church of Jesus Christ of Latter Day Saints, a community which practices both endogamy and polygyny, where an estimated 75 to 80% of the community are blood relatives of just two men—founders John Y. Barlow and Joseph Smith Jessop.[26]
Around 1814, a small group of British colonists founded a settlement on Tristan da Cunha, a group of small islands in the Atlantic Ocean, midway between Africa and South America. One of the early colonists apparently carried a rare, recessive allele for retinitis pigmentosa, a progressive form of blindness that afflicts homozygous individuals. As late as 1961, the majority of the genes in the gene pool on Tristan were still derived from 15 original ancestors; as a consequence of the inbreeding, of 232 people tested in 1961, fully four were suffering from retinitis pigmentosa.[27]
More severe illnesses exist among certain Jewish groups. Ashkenazi Jews, for example, have a particularly high chance of suffering from Tay-Sachs disease, a fatal condition in young children (see Medical genetics of Ashkenazi Jews). Ashkenazi Jews display a strong genetic bottleneck. (a similar phenomena to the founder effect)
The abnormally high rate of twin births in Cândido Godói could be explained by the founder effect.[28]
Founder effects in language
The founder effect in language underlines the impact of the founder population on the nature of a language that emerges from reclamation or creolization. For example, according to linguist Ghil'ad Zuckermann, "Yiddish is a primary contributor to Israeli Hebrew because it was the mother tongue of the vast majority of revivalists and first pioneers in Eretz Yisrael at the crucial period of the beginning of Israeli Hebrew".[29] According to Zuckermann, although the Hebrew revivalists (i.e. the founders of the Israeli language) wished to speak Hebrew, with Semitic grammar and pronunciation, they could not avoid the Ashkenazi mindset deriving from their European background. He argues that the revivalists' attempt to deny their European roots, negate diasporism, and avoid hybridity as reflected in Yiddish, failed. "Had the revivalists been Arabic-speaking Jews (e.g. from Morocco), Israeli Hebrew would have been a totally different language – both genetically and typologically, much more Semitic."[29] Such linguistic "Founder Principle" led Zuckermann to suggest that Israeli (the language spoken in Israel today) is a Semito-European hybrid rather than merely Hebrew.
See also
References
- ↑ Provine, W. B. (2004). "Ernst Mayr: Genetics and speciation". Genetics. 167 (3): 1041–6. PMC 1470966
 . PMID 15280221.
. PMID 15280221. - ↑ Templeton, A. R. (1980). "The theory of speciation via the founder principle". Genetics. 94 (4): 1011–38. PMC 1214177. PMID 6777243.
- ↑ Hartwell, Leland; Hood, Leroy; Goldberg, Michael; Reynolds, Ann E.; Silver, Lee; Veres, Ruth C (2004). Genetics: From Genes to Genomes. p. 241. ISBN 978-0-07-121468-1.
- ↑ Raven, Peter H.; Evert, Ray F.; Eichhorn, Susan E. (1999). Biology of Plants. W H Freeman and Company. p. 241.
- ↑ "Bioinformatics Glossary". bscs.org. Archived from the original on March 25, 2009. Retrieved 2009-03-23.
- ↑ "Colorectal Cancer Research Definitions". www.mshri.on.ca. Archived from the original on July 24, 2009. Retrieved 2009-03-23.
- ↑ "Founder Mutations: Scientific American". www.sciam.com. Retrieved 2009-03-23.
- ↑ Wade, Michael S.; Wolf, Jason; Brodie, Edmund D. (2000). Epistasis and the evolutionary process. Oxford [Oxfordshire]: Oxford University Press. p. 330. ISBN 0-19-512806-0.
- ↑ Mayr, Ernst, Jody Hey, Walter M. Fitch, Francisco José Ayala (2005). Systematics and the Origin of Species: on Ernst Mayr's 100th anniversary (Illustrated ed.). National Academies Press. p. 367. ISBN 978-0-309-09536-5.
- ↑ http://evolution.berkeley.edu/evosite/evo101/VC1cPeripatric.shtml[]
- ↑ Howard, Daniel J.; Berlocher, Steward H. (1998). Endless Forms (Illustrated ed.). United States: Oxford University Press. p. 470. ISBN 978-0-19-510901-6.
- ↑ Ramachandran, S.; Deshpande, O.; Roseman, C. C.; Rosenberg, N. A.; Feldman, M. W.; Cavalli-Sforza, L. L. (2005). "Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa". Proceedings of the National Academy of Sciences. 102 (44): 15942–7. Bibcode:2005PNAS..10215942R. doi:10.1073/pnas.0507611102. JSTOR 4143304. PMC 1276087. PMID 16243969.
- ↑ Degiorgio, M.; Jakobsson, M.; Rosenberg, N. A. (2009). "Explaining worldwide patterns of human genetic variation using a coalescent-based serial founder model of migration outward from Africa". Proceedings of the National Academy of Sciences. 106 (38): 16057–62. Bibcode:2009PNAS..10616057D. doi:10.1073/pnas.0903341106. JSTOR 40485019. PMC 2752555. PMID 19706453.
- ↑ Maji, Suvendu; Krithika, S.; Vasulu, T. S. (2009). "Phylogeographic distribution of mitochondrial DNA macrohaplogroup M in India". Journal of Genetics. 88 (1): 127–39. doi:10.1007/s12041-009-0020-3. PMID 19417557.
- ↑ Thangaraj, Kumarasamy; Chaubey, Gyaneshwer; Singh, Vijay; Vanniarajan, Ayyasamy; Thanseem, Ismail; Reddy, Alla G; Singh, Lalji (2006). "In situ origin of deep rooting lineages of mitochondrial Macrohaplogroup 'M' in India". BMC Genomics. 7: 151. doi:10.1186/1471-2164-7-151. PMC 1534032. PMID 16776823.
- ↑ Hajji, Ghaiet M.; Charfi-Cheikrouha, F.; Lorenzini, Rita; Vigne, Jean-Denis; Hartl, Günther B.; Zachos, Frank E. (2007). "Phylogeography and founder effect of the endangered Corsican red deer (Cervus elaphus corsicanus)". Biodiversity and Conservation. 17 (3): 659–73. doi:10.1007/s10531-007-9297-9.
- ↑ Hundertmark, Kris J.; Van Daele, Larry J. (2009). "Founder effect and bottleneck signatures in an introduced, insular population of elk". Conservation Genetics. 11: 139–47. doi:10.1007/s10592-009-0013-z.
- ↑ Kolbe, J. J.; Leal, M.; Schoener, T. W.; Spiller, D. A.; Losos, J. B. (2012). "Founder Effects Persist Despite Adaptive Differentiation: A Field Experiment with Lizards". Science. 335 (6072): 1086–9. Bibcode:2012Sci...335.1086K. doi:10.1126/science.1209566. PMID 22300849.
- ↑ Charbonneau, Hubert; Desjardins, Bertrand; Légaré, Jacques; Denis, Hubert (2010). "The Population of the St. Lawrence Valley 1608–1760". In Haines, Michael R.; Stecke, Richard H. A Population History of North America. pp. 99–142. ISBN 978-0-521-49666-7.
- ↑ Bherer, Claude; Labuda, Damian; Roy-Gagnon, Marie-Hélène; Houde, Louis; Tremblay, Marc; Vézina, Hélène (2011). "Admixed ancestry and stratification of Quebec regional populations". American Journal of Physical Anthropology. 144 (3): 432–41. doi:10.1002/ajpa.21424. PMID 21302269.
- ↑ McKusick, V. A.; Egeland, J. A.; Eldridge, R; Krusen, D. E. (1964). "Dwarfism in the Amish I. The Ellis-Van Creveld Syndrome". Bulletin of the Johns Hopkins Hospital. 115: 306–36. PMID 14217223.
- 1 2 http://rarediseases.about.com/od/rarediseases1/a/062004.htm
About.com: Rare diseases - June 2004. Maple syrup urine disease by Mary Kugler, R.N.
Article describes MSUD prevalence among Amish and Mennonite children. - ↑ Jaworski, M. A.; Severini, A; Mansour, G; Konrad, H. M.; Slater, J; Hennig, K; Schlaut, J; Yoon, J. W.; Pak, C. Y.; MacLaren, N (1989). "Genetic conditions among Canadian Mennonites: Evidence for a founder effect among the old colony (Chortitza) Mennonites". Clinical and investigative medicine. Medecine clinique et experimentale. 12 (2): 127–41. PMID 2706837.
- ↑ Puffenberger, E.G. (2003). "Genetic heritage of the Old Order Mennonites of southeastern Pennsylvania". American Journal of Medical Genetics. 121C (1): 18–31. doi:10.1002/ajmg.c.20003. PMID 12888983.
- ↑ http://www.mazornet.com/genetics/MapleSyrup.htm[]
- ↑ Forbidden Fruit:Inbreeding among polygamists along the Arizona-Utah border is producing a caste of severely retarded and deformed children, by John Dougherty, The Phoenix New Times News, December 29, 2005, page 2.
- ↑ Berry, RJ (1967). "Genetical changes in mice and men" (PDF). Eugen Rev. 59: 78–96. PMC 2906351. PMID 4864588.
- ↑ De Oliveira, Marcelo Zagonel; Schüler-Faccini, Lavínia; Demarchi, Dario A.; Alfaro, Emma L.; Dipierri, José E.; Veronez, Mauricio R.; Colling Cassel, Marlise; Tagliani-Ribeiro, Alice; Silveira Matte, Ursula; Ramallo, Virginia (2013). "So Close, So Far Away: Analysis of Surnames in a Town of Twins (Cândido Godói, Brazil)". Annals of Human Genetics. 77 (2): 125–36. doi:10.1111/ahg.12001. PMID 23369099.
- 1 2 See p. 63 in Zuckermann, Ghil'ad (2006). "A New Vision for 'Israeli Hebrew': Theoretical and Practical Implications of Analysing Israel's Main Language as a Semi-Engineered Semito-European Hybrid Language" (PDF). Journal of Modern Jewish Studies. 5 (1): 57–71. doi:10.1080/14725880500511175. Retrieved September 9, 2016.
Further reading
- Mayr, Ernst (1954). "Change of genetic environment and evolution". In Julian Huxley. Evolution as a Process. London: George Allen & Unwin. OCLC 974739.
- Mayr, Ernst (1963). Animal Species and Evolution. Cambridge: Belknap Press of Harvard University Press. ISBN 0-674-03750-2.
External links
| Wikimedia Commons has media related to Founder effect. |