Group I catalytic intron
| Group I catalytic intron | |
|---|---|
 | |
| Predicted secondary structure and sequence conservation of Group I catalytic intron | |
| Identifiers | |
| Symbol | Intron_gpI |
| Rfam | RF00028 |
| Other data | |
| RNA type | Intron |
| Domain(s) | Eukaryota; Bacteria; Viruses |
| GO | 0000372 |
| SO | 0000587 |
Group I introns are large self-splicing ribozymes. They catalyze their own excision from mRNA, tRNA and rRNA precursors in a wide range of organisms.[1][2][3] The core secondary structure consists of nine paired regions (P1-P9).[4] These fold to essentially two domains - the P4-P6 domain (formed from the stacking of P5, P4, P6 and P6a helices) and the P3-P9 domain (formed from the P8, P3, P7 and P9 helices).[2] The secondary structure mark-up for this family represents only this conserved core. Group I introns often have long open reading frames inserted in loop regions.
Catalysis
Splicing of group I introns is processed by two sequential ester-transfer reactions.[3] The exogenous guanosine or guanosine nucleotide (exoG) first docks onto the active G-binding site located in P7, and its 3'-OH is aligned to attack the phosphodiester bond at the 5' splice site located in P1, resulting in a free 3'-OH group at the upstream exon and the exoG being attached to the 5' end of the intron. Then the terminal G (omega G) of the intron swaps the exoG and occupies the G-binding site to organize the second ester-transfer reaction: the 3'-OH group of the upstream exon in P1 is aligned to attack the 3' splice site in P10, leading to the ligation of the adjacent upstream and downstream exons and release of the catalytic intron.
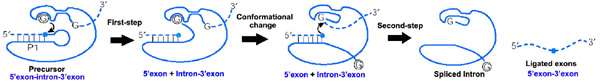
Two-metal-ion mechanism seen in protein polymerases and phosphatases was proposed to be used by group I and group II introns to process the phosphoryl transfer reactions,[5] which was unambiguously proven by a recently resolved high-resolution structure of the Azoarcus group I intron.[6]

Intron folding
Since the early 1990s, scientists started to study how the group I intron achieves its native structure in vitro, and some mechanisms of RNA folding have been appreciated thus far.[10] It is agreed that the tertiary structure is folded after the formation of the secondary structure. During folding, RNA molecules are rapidly populated into different folding intermediates, the intermediates containing native interactions are further folded into the native structure through a fast folding pathway, while those containing non-native interactions are trapped in metastable or stable non-native conformations, and the process of conversion to the native structure occurs very slowly. It is evident that group I introns differing in the set of peripheral elements display different potentials in entering the fast folding pathway. Meanwhile, cooperative assembly of the tertiary structure is important for the folding of the native structure. Nevertheless, folding of group I introns in vitro encounters both thermodynamic and kinetic challenges. A few RNA binding proteins and chaperones have been shown to promote the folding of group I introns in vitro and in bacteria by stabilizing the native intermediates, and by destabilizing the non-native structures, respectively.
Distribution, phylogeny and mobility
Group I introns are distributed in bacteria, lower eukaryotes and higher plants. However, their occurrence in bacteria seems to be more sporadic than in lower eukaryotes, and they have become prevalent in higher plants. The genes that group I introns interrupt differ significantly: They interrupt rRNA, mRNA and tRNA genes in bacterial genomes, as well as in mitochondrial and chloroplast genomes of lower eukaryotes, but only invade rRNA genes in the nuclear genome of lower eukaryotes. In higher plants, these introns seem to be restricted to a few tRNA and mRNA genes of the chloroplasts and mitochondria.
Group I introns are also found inserted into genes of a wide variety of bacteriophages of Gram-positive bacteria.[11] However, their distribution in the phage of Gram-negative bacteria is mainly limited to the T4, T-even and T7-like bacteriophages.[11][12][13][14]
Both intron-early and intron-late theories have found evidences in explaining the origin of group I introns. Some group I introns encode homing endonuclease (HEG), which catalyzes intron mobility. It is proposed that HEGs move the intron from one location to another, from one organism to another and thus account for the wide spreading of the selfish group I introns. No biological role has been identified for group I introns thus far except for splicing of themselves from the precursor to prevent the death of the host that they live by. A small number of group I introns are also found to encode a class of proteins called maturases that facilitate the intron splicing.
See also
- Intron
- Group I Intron Sequence and Structure Database
- Splice site
- Nuclear introns
- Group II intron
- Group III intron
- Twintron
- LtrA
- Cyclic di-GMP-II riboswitch, where there is an example of a riboswitch acting together with a group I intron to regulate the expression of a gene
References
- ↑ Nielsen H, Johansen SD (2009). "Group I introns: Moving in new directions". RNA Biol. 6 (4): 375–83. doi:10.4161/rna.6.4.9334. PMID 19667762. Retrieved 2010-07-15.
- 1 2 Cate JH, Gooding AR, Podell E, et al. (September 1996). "Crystal structure of a group I ribozyme domain: principles of RNA packing". Science. 273 (5282): 1678–85. doi:10.1126/science.273.5282.1678. PMID 8781224. Retrieved 2010-07-15.
- 1 2 Cech TR (1990). "Self-splicing of group I introns". Annu. Rev. Biochem. 59: 543–68. doi:10.1146/annurev.bi.59.070190.002551. PMID 2197983. Retrieved 2010-07-15.
- ↑ Woodson SA (June 2005). "Structure and assembly of group I introns". Curr. Opin. Struct. Biol. 15 (3): 324–30. doi:10.1016/j.sbi.2005.05.007. PMID 15922592. Retrieved 2010-07-15.
- ↑ Steitz, TA; Steitz JA (1993). "A general two-metal-ion mechanism for catalytic RNA". Proc Natl Acad Sci USA. 90 (14): 6498–6502. doi:10.1073/pnas.90.14.6498. PMC 46959
 . PMID 8341661.
. PMID 8341661. - ↑ Stahley, MR; Strobel SA (2006). "RNA splicing: group I intron crystal structures reveal the basis of splice site selection and metal ion catalysis". Curr Opin Struct Biol. 16 (3): 319–326. doi:10.1016/j.sbi.2006.04.005. PMID 16697179.
- ↑ Golden BL, Gooding AR, Podell ER, Cech TR (1998). "A preorganized active site in the crystal structure of the Tetrahymena ribozyme". Science. 282 (5387): 259–64. doi:10.1126/science.282.5387.259. PMID 9841391.
- ↑ Golden BL, Kim H, Chase E (2005). "Crystal structure of a phage Twort group I ribozyme-product complex". Nat Struct Mol Biol. 12 (1): 82–9. doi:10.1038/nsmb868. PMID 15580277.
- ↑ Guo F, Gooding AR, Cech TR (2004). "Structure of the Tetrahymena ribozyme: base triple sandwich and metal ion at the active site". Mol Cell. 16 (3): 351–62. doi:10.1016/j.molcel.2004.10.003. PMID 15525509.
- ↑ Brion P, Westhof E (1997). "Hierarchy and dynamics of RNA folding". Annu Rev Biophys Biomol Struct. 26: 113–37. doi:10.1146/annurev.biophys.26.1.113. PMID 9241415. Retrieved 2010-07-15.
- 1 2 Edgell DR, Belfort M, Shub DA (October 2000). "Barriers to intron promiscuity in bacteria". J. Bacteriol. 182 (19): 5281–9. doi:10.1128/jb.182.19.5281-5289.2000. PMC 110968. PMID 10986228.
- ↑ Sandegren L, Sjöberg BM (May 2004). "Distribution, sequence homology, and homing of group I introns among T-even-like bacteriophages: evidence for recent transfer of old introns". J. Biol. Chem. 279 (21): 22218–27. doi:10.1074/jbc.M400929200. PMID 15026408.
- ↑ Bonocora RP, Shub DA (December 2004). "A self-splicing group I intron in DNA polymerase genes of T7-like bacteriophages". J. Bacteriol. 186 (23): 8153–5. doi:10.1128/JB.186.23.8153-8155.2004. PMC 529087. PMID 15547290.
- ↑ Lee CN, Lin JW, Weng SF, Tseng YH (December 2009). "Genomic characterization of the intron-containing T7-like phage phiL7 of Xanthomonas campestris". Appl. Environ. Microbiol. 75 (24): 7828–37. doi:10.1128/AEM.01214-09. PMC 2794104. PMID 19854925.
Further reading
- Chauhan, S; Caliskan G; Briber RM; Perez-Salas U; Rangan P; Thirumalai D; Woodson SA (2005). "RNA tertiary interactions mediate native collapse of a bacterial group I ribozyme". J Mol Biol. 353 (5): 1199–1209. doi:10.1016/j.jmb.2005.09.015. PMID 16214167.
- Haugen, P; Simon DM; Bhattacharya D (2005). "The natural history of group I introns". TRENDS in Genetics. 21 (2): 111–119. doi:10.1016/j.tig.2004.12.007. PMID 15661357.
- Rangan, P; Masquida, B; Westhof E; Woodson SA (2003). "Assembly of core helices and rapid tertiary folding of a small bacterial group I ribozyme". Proc Natl Acad Sci USA. 100 (4): 1574–1579. doi:10.1073/pnas.0337743100. PMC 149874. PMID 12574513.
- Schroeder, R; Barta A; Semrad K (2004). "Strategies for RNA folding and assembly". Nat Rev Mol Cell Biol. 5 (11): 908–919. doi:10.1038/nrm1497. PMID 15520810.
- Thirumalai, D; Lee N; Woodson SA; Klimov D (2001). "Early events in RNA folding". Annu Rev Phys Chem. 52: 751–762. doi:10.1146/annurev.physchem.52.1.751. PMID 11326079.
- Treiber, DK; Williamson JR (1999). "Exposing the kinetic traps in RNA folding". Curr Opin Struct Biol. 9 (3): 339–345. doi:10.1016/S0959-440X(99)80045-1. PMID 10361090.
- Xiao, M; Leibowitz MJ; Zhang Y (2003). "Concerted folding of a Candida ribozyme into the catalytically active structure posterior to a rapid RNA compaction". Nucleic Acids Res. 31 (14): 3901–3908. doi:10.1093/nar/gkg455. PMC 165970. PMID 12853605.