Temperature-dependent sex determination
Temperature-dependent sex determination (TSD) is a type of environmental sex determination in which the temperatures experienced during embryonic development determine the sex of the offspring.[1] It is most prevalent and common among amniote vertebrates that are classified under the reptile class.[2] TSD differs from the chromosomal sex-determination systems common among vertebrates. It is a type of environmental sex determination (ESD); in other ESD systems, some factors such as population determine the sex of organisms (see Polyphenism). TSD was also thought to occur in some megapodes, such as the Australian brushturkey; however, their offspring sex ratios appear to result from temperature-dependent embryo mortality rather than from TSD.[3]
The eggs are affected by the temperature at which they are incubated during the middle one-third of embryonic development.[4] This critical period of incubation is known as the thermosensitive period (TSP).[5] The specific time of sex-commitment is known due to several authors resolving histological chronology of sex differentiation in the gonads of turtles with TSD.[4]
Thermosensitive Period (TSP)
The thermosensitive, or temperature-sensitive, period (TSP) is the period during development when sex is irreversibly determinated. It is used in reference to species with temperature-dependent sex determination, such as alligators and turtles.[6] The TSP typically spans the middle third of incubation with the endpoints defined by embryonic stage. The extent of the TSP varies a little among species,[6] and development within the oviducts must be taken into account in species where the embryo is at a relatively late stage of development on egglaying (e.g. many lizards). Temperature pulses during the thermosensitive period are often sufficient to determine sex, but after the TSP, sex is unresponsive to temperature. After this period, however, sex cannot be reversed (see sex reversal).[6]
Types
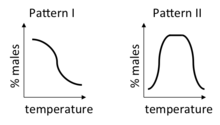
Within the mechanism, two distinct patterns have been discovered and named Pattern I and Pattern II, with Pattern I further divided into IA and IB. Pattern IA has a single transition zone, where eggs predominantly hatch males if incubated below this temperature zone, and predominantly hatch females if incubated above it. Pattern 1A occurs in most turtles, with the transition between male-producing temperatures and female-producing temperatures occurring over a range of temperatures as little as 1–2 °C.[8] Pattern IB also has a single transition zone, but females are produced below it and males above it. Pattern 1B occurs in the Tuatara. Pattern II has two transition zones, with males dominating at intermediate temperatures and females dominating at both extremes.[9] Pattern II occurs in some turtles, lizards and crocodilians. Very near or at the pivotal temperature of sex determination, mixed sex ratios and, more rarely, intersex individuals are produced.[8]
It has been proposed that essentially all modes of TSD are actually type II and those that deviate from the expected female-male-female pattern are simply never exposed to extreme temperature ranges on one end of the range or the other.[10]
The distinction between chromosomal sex-determination systems and TSD is often blurred, because the sex of some species, such as the three-lined skink Bassiana duperreyi and the central bearded dragon Pogona vitticeps, is determined by sex chromosomes, but this is over-ridden by temperatures that are tolerable but extreme. Also, experiments conducted at the pivotal temperature, where temperature is equivocal in its influence, have demonstrated an underlying genetic predisposition to be one sex or the other.
Examples
Temperature-dependent sex determination was first described in Agama agama in the year 1966 by Madeleine Charnier.[11]
A 2015 study found that hot temperatures altered the expression of the sex chromosomes in Australia's bearded dragon lizards. The lizards were female in appearance, and were capable of bearing offspring, despite having the ZZ chromosomes usually associated with male lizards.[12]
Hormones in TSD systems
Synergism between temperature and hormones has also been identified in these systems. Administering estradiol at male-producing temperatures generates females that are physiologically identical to temperature-produced females.[13] The reverse experiment, males produced at female temperatures, only occurs when a nonaromatizable testosterone or an aromatase inhibitor is administered, indicating that the enzyme responsible for conversion of testosterone to estradiol, aromatase, plays a role in female development.[14] Nonetheless, the mechanisms for TSD are still relatively unknown, but in some ways, TSD resembles genetic sex determination (GSD), particularly in regards to the effects of aromatase in each process.[15] In some fish species, aromatase is in both the ovaries of female organisms who underwent TSD and those who underwent GSD, with no less than 85% of the coding sequences of each aromatase being identical,[16] showing that aromatase is not unique to TSD and suggesting that there must be another factor in addition to it that is also affecting TSD.
Interestingly, hormones and temperature show signs of acting in the same pathway, in that less hormone is required to produce a sexual shift as the incubation conditions near the pivotal temperature. It has been proposed[17] that temperature acts on genes coding for such steroidogenic enzymes, and testing of homologous GSD pathways has provided a genic starting point.[18] Yet, the genetic sexual determination pathway in TSD turtles is poorly understood and the controlling mechanism for male or female commitment has not been identified.[19]
While sex hormones have been observed to be influenced by temperature, thus potentially altering sexual phenotypes, specific genes in the gonadal differentiation pathway display temperature influenced expression.[20] In some species such important sex-determining genes as DMRT1[21] and those involved in the Wnt signalling pathway [20] could potentially be implicated as genes which provide a mechanism (opening the door for selective forces) for the evolutionary development of TSD. While aromatase is involved in more processes than only TSD, it has also been shown to play a role in certain tumor development.[22]
Adaptive significance
The adaptive significance of TSD is currently not well understood. One possible explanation that TSD is common in amniotes is phylogenetic inertia – TSD is the ancestral condition in this clade and is simply maintained in extant lineages because it is currently adaptively neutral or nearly so.[23] Indeed, recent phylogenetic comparative analyses imply a single origin for TSD in most amniotes around 300 million years, with the re-evolution of TSD in squamates [24] and turtles[25] after they had independently developed GSD. Consequently, the adaptive significance of TSD in all but the most recent origins of TSD may have been obscured by the passage of deep time, with TSD potentially being maintained in many amniote clades simply because it works ‘well enough’ (i.e. has no overall fitness costs along the lines of the phylogenetic inertia explanation).
Other work centers on a 1977 theoretical model (the Charnov–Bull model),[26][27] predicted that selection should favour TSD over chromosome-based systems when "the developmental environment differentially influences male versus female fitness";[2] this theoretical model was empirically validated thirty years later[2] but the generality of this hypothesis in reptiles is questioned. This hypothesis is supported by the persistence of TSD in certain populations of spotted skink (Niveoscincus ocellatus), a small lizard in Tasmania, where it is advantageous to have females early in the season. The warmth early in the season ensures female-biased broods that then have more time to grow and reach maturity and possibly reproduce before they experience their first winter, thereby increasing fitness of the individual.[1]
In support of the Charnov and Bull hypothesis, Warner and Shine (2008) showed confidently that incubation temperature influences males’ reproductive success differently than females in Jacky Dragon lizards (Amphibolurus muricatus) by treating the eggs with chemicals that interfere with steroid hormone biosynthesis. These chemicals block the conversion of testosterone to oestradiol during development so each sex offspring can be produced at all temperatures. They found that hatching temperatures that naturally produce each sex maximized fitness of each sex, which provides the substantial empirical evidence in support of the Charnov & Bull model for reptiles.[28]
Spencer and Janzen (2014) found further support for the Charnov-Bull model by incubating painted turtles (Chrysemys picta) at different temperatures and measuring various characteristics indicative of fitness. The turtles were incubated at temperatures that produce solely males, both sexes, and solely females. Spencer and Janzen (2014) found that hatchlings from mixed-sex nests were less energy efficient and grew less than their same-sex counterparts incubated in single-sex producing temperatures. Hatchlings from single-sex producing temperatures also had higher first-year survivorship than the hatchlings from the temperature that produces both sexes. TSD may be advantageous and selected for in turtles, as embryo energy efficiency and hatchling size are optimized for each sex at single-sex incubation temperatures and are indicative of first-year survivorship.[29] This suggests that natural selection would favor TSD, as TSD may enhance the fitness of offspring.
An alternative hypothesis of adaptive significance was proposed by Bulmer and Bull in 1982[30] and supported by the work of Pen et al. (2010). They conjectured that disruptive selection produced by variation in the environment could result in an evolutionary transition from ESD to GSD (Bull, Vogt, and Bulmer, 1982). Pen et al. (2010) addresses evolutionary divergence in SDM’s via natural selection on sex ratios. Studying the spotted skink, they observed that the highland population was not affected by temperature, yet there was a negative correlation between annual temperature and cohort sex ratios in the lowlands. The highlands are colder with a higher magnitude of annual temperature fluctuation and a shorter activity season, delaying maturity, thus GSD is favored so sex ratios are not skewed. However, in the lowlands, temperatures are more constant and a longer activity season allows for favorable conditions for TSD. They concluded that this differentiation in climate causes divergent selection on regulatory elements in the sex-determining network allowing for the emergence of sex chromosomes in the highlands.[31]
"Temperature sex determination could allow the mother to determine the sex of her offspring by varying the temperature of the nest in which her eggs are incubated. However there is no evidence thus far that sex ratio is manipulated by parental care." [32]
The effect of climate change
The warming of the habitats of species exhibiting TSD are beginning to affect their behavior and may soon start affecting their physiology.[33] Many species (Pattern IA and II) have begun to nest earlier and earlier in the year to preserve the sex ratio.[34] The three traits of pivotal temperature (the temperature at which the sex ratio is 50%), maternal nest-site choice, and nesting phenology have been identified as the key traits of TSD that can change, and of these, only the pivotal temperature is significantly heritable, and unfortunately this would have to increase by 27 standard deviations to compensate for a 4 °C temperature increase.[35] It is likely that climate change will outpace the ability of many animals to adapt,[36][37] and many will likely go extinct. However, there is evidence that during climactic extremes, changes in the sex determining mechanism itself (to GSD) are selected for, particularly in the highly-mutable turtles.[38]
See also
References
- 1 2 Pen, Ido, Tobias Uller, Barbara Feldmeyer, Anna Harts, Geoffrey M. While, and Erik Wapstra (2010). "Climate-driven population divergence in sex-determining systems". Nature. 468: 436–439. doi:10.1038/nature09512.
- 1 2 3 Warner DA, Shine R (2008). "The adaptive significance of temperature-dependent sex determination in a reptile". Nature. 451 (7178): 566–568. doi:10.1038/nature06519. PMID 18204437.
- ↑ Göth, Ann; Booth, David T (22 March 2005). "Temperature-dependent sex ratio in a bird". Biology Letters. 1: 31–33. doi:10.1098/rsbl.2004.0247. PMC 1629050
 . PMID 17148121.
. PMID 17148121. - 1 2 Wibbels, T.; Bull, J.J.; Crews, D. (1991). "Chronology and morphology of temperature dependent sex determination". The Journal of Experimental Zoology. 260 (3): 371–381. doi:10.1002/jez.1402600311. PMID 1744617.
- ↑ Delmas, V.; Prevot-Julliard, A.-C.; Pieau, C.; Girondot, M. (2008). "A mechanistic model of temperature-dependent sex determination in a chelonian: the European pond turtle". Functional Ecology. 22: 84–93. doi:10.1111/j.1365-2435.2007.01349.x.
- 1 2 3 Lance, V. A. (2009). "Is regulation of aromatase expression in reptiles the key to understanding temperature-dependent sex determination?". Journal of Experimental Zoology Part A: Ecological Genetics and Physiology. 311A (5): 314–322. doi:10.1002/jez.465. PMID 18668631.
- ↑ Shoemaker, C. M.; Crews, D (2009). "Analyzing the coordinated gene network underlying temperature-dependent sex determination in reptiles". Seminars in Cell & Developmental Biology. 20 (3): 293–303. doi:10.1016/j.semcdb.2008.10.010. PMC 2729108. PMID 19022389.
- 1 2 Bull, J. J. (1980). "Sex determination in reptiles". Quart. Review of Biology. 55: 3–21. doi:10.1086/411613. JSTOR 2826077.
- ↑ Eti, turtles of the world. "Temperature dependent sex determination". ETI. Accessed April 16, 2008
- ↑ Graves, J.A.M. (2008). "Weird animal genomes and the evolution of vertebrate sex and sex chromosomes.". Annual Review of Genetics. 42: 565–86. doi:10.1146/annurev.genet.42.110807.091714. PMID 18983263.
- ↑ https://embryo.asu.edu/pages/temperature-dependent-sex-determination-reptiles
- ↑ NBC (1 July 2015). "Hotter Temperatures Trigger Sex Change in Australian Lizards".
- ↑ Wibbels, T.; Bull, J.J.; Crews, D. (1991). "Synergism between temperature and estradiol: A common pathway in turtle sex determination". The Journal of Experimental Zoology. 260 (1): 130–134. doi:10.1002/jez.1402600117. PMID 1791420.
- ↑ Crews, D (1996). "Temperature-dependent sex determination: the interplay of steroid hormones and temperature.". Zoo. Sci. 13 (1): 1–13. doi:10.2108/zsj.13.1. PMID 8688803.
- ↑ Kohno, S.; Katsu, Y.; Urushitani, H.; Ohta, Y.; Iguchi, T.; Guillette, J. (2010). "Potential Contributions of Heat Shock Proteins to Temperature-Dependent Sex Determination in the American Alligator". Sexual Development. 4 (1–2): 73–87. doi:10.1159/000260374. PMC 2855287. PMID 19940440.
- ↑ Duffy, T. A.; Picha, M. E.; Won, E. T.; Borski, R. J.; McElroy, A. E.; Conover, D. O. (2010). "Ontogenesis of gonadal aromatase gene expression in atlantic silverside (Menidia menidia) Populations with genetic and temperature-dependent sex determination". Journal of Experimental Zoology Part A: Ecological Genetics and Physiology. 313A (7): 421–431. doi:10.1002/jez.612. PMID 20623799.
- ↑ Crews, D. (2003). "Sex determination: where environment and genetics meet". Evolution and Development. 5 (1): 50–55. doi:10.1046/j.1525-142X.2003.03008.x. PMID 12492409.
- ↑ Miller, D.; Summers, J.; Silber, S. (2004). "Environmental versus genetic sex determination: a possible factor in dinosaur extinction?". Fertility and Sterility. 81 (4): 954–964. doi:10.1016/j.fertnstert.2003.09.051. PMID 15066448.
- ↑ Georges, A.; Ezaz, T.; Quinn, A. E.; Sarre, S. D. (2010). "Are reptiles predisposed to temperature-dependent sex determination?". Sexual Development. 4 (1–2): 7–15. doi:10.1159/000279441. PMID 20110654.
- 1 2 Valenzuela, N. (2008). "Evolution of the gene network underlying gonadogenesis in turtles with temperature-dependent and genotypic sex determination". Integrative and Comparative Biology. 48 (4): 476–485. doi:10.1093/icb/icn031. PMID 21669808.
- ↑ Graves, J.A.M. (2009). "Weird animal genomes and the evolution of human sex chromosomes". Hormone Research. 72: 15–15.
- ↑ Akinci, H.; Kapucu, A.; Dar, K. A.; Celik, O.; Tutunculer, B.; Sirin, G.; Oz, B.; Gazioglu, N.; Ince, H.; Aliustaoglu, S. H.; Kadioglu, P. (2012). "Aromatase cytochrome P450 enzyme expression in prolactinomas and its relationship to tumor behavior". Pituitary. 16 (3): 386–392. doi:10.1007/s11102-012-0436-2. PMID 22983691.
- ↑ Janzen, F. J.; Phillips, P. C. (2006). "Exploring the evolution of environmental sex determination, especially in reptiles". Journal of Evolutionary Biology. 19 (6): 1775–1784. doi:10.1111/j.1420-9101.2006.01138.x. PMID 17040374.
- ↑ Janzen & Krenz, 2004
- ↑ Valenzuela, Nicole & Dean C. Adams (2011). "Chromosome number and sex determination coevolve in turtles". Evolution. 65 (6): 1808–1813. doi:10.1111/j.1558-5646.2011.01258.x. PMID 21644965.
- ↑ Bull JJ, Charnov EL (1977). "Changes in the heterogametic mechanism of sex determination". Heredity. 39 (1): 1–14. doi:10.1038/hdy.1977.38. PMID 268319.
- ↑ Charnov EL, Bull J (1977). "When is sex environmentally determined?". Nature. 266 (5605): 828–830. doi:10.1038/266828a0. PMID 865602.
- ↑ Warner, D.A; Shine, R (Jan 31, 2008). "The adaptive significance of temperature-dependent sex determination in a reptile". Nature. 451 (7178): 566–U5. doi:10.1038/nature06519. PMID 18204437.
- ↑ Spencer, R. J.; Janzen, F. J. (2014). "A novel hypothesis for the adaptive maintenance of environmental sex determination in a turtle". Proceedings of the Royal Society B. 281: 20140831. doi:10.1098/rspb.2014.0831.
- ↑ Bull, J.J.; Vogt, R.C.; Bulmer, M.G. (1982). "Heritability of Sex Ratio in Turtles with Environmental Sex Determination". Evolution. 36 (2): 333–341. doi:10.2307/2408052.
- ↑ Pen, Ido; Uller, Tobias; Feldmeyer, Barbara; Harts, Anna; While, Geoffrey M.; Wapstra, Erik (Nov 18, 2010). "Climate-driven population divergence in sex-determining systems". Nature. 468 (7322): 436–U262. doi:10.1038/nature09512. PMID 20981009.
- ↑ Nelson, Randy. An Introduction to Behavioral Endocrinology. Sinauer Associates: Massachusetts. 2005. pg 136
- ↑ Schwanz, Lisa E.; Janzen, Fredric J. (2008-11-01). "Climate Change and Temperature‐Dependent Sex Determination: Can Individual Plasticity in Nesting Phenology Prevent Extreme Sex Ratios?". Physiological and Biochemical Zoology. 81 (6): 826–834. doi:10.1086/590220. JSTOR 10.1086/590220.
- ↑ Hawkes, L. A.; Broderick, A. C.; Godfrey, M. H.; Godley, B. J. (2007-05-01). "Investigating the potential impacts of climate change on a marine turtle population". Global Change Biology. 13 (5): 923–932. doi:10.1111/j.1365-2486.2007.01320.x. ISSN 1365-2486.
- ↑ Refsnider, Jeanine M.; Janzen, Fredric J. (2015-08-05). "Temperature-Dependent Sex Determination under Rapid Anthropogenic Environmental Change: Evolution at a Turtle's Pace?". Journal of Heredity. 107: esv053. doi:10.1093/jhered/esv053. ISSN 0022-1503. PMID 26245920.
- ↑ Hays, Graeme C.; Fossette, Sabrina; Katselidis, Kostas A.; Schofield, Gail; Gravenor, Mike B. (2010-12-01). "Breeding Periodicity for Male Sea Turtles, Operational Sex Ratios, and Implications in the Face of Climate Change". Conservation Biology. 24 (6): 1636–1643. doi:10.1111/j.1523-1739.2010.01531.x. ISSN 1523-1739.
- ↑ Santidrián Tomillo, Pilar; Genovart, Meritxell; Paladino, Frank V.; Spotila, James R.; Oro, Daniel (2015-08-01). "Climate change overruns resilience conferred by temperature-dependent sex determination in sea turtles and threatens their survival". Global Change Biology. 21 (8): 2980–2988. doi:10.1111/gcb.12918. ISSN 1365-2486.
- ↑ Valenzuela, Nicole; Adams, Dean C. (2011-06-01). "Chromosome Number and Sex Determination Coevolve in Turtles". Evolution. 65 (6): 1808–1813. doi:10.1111/j.1558-5646.2011.01258.x. ISSN 1558-5646. PMID 21644965.