Suillellus luridus
| Suillellus luridus | |
|---|---|
 | |
| Scientific classification | |
| Kingdom: | Fungi |
| Division: | Basidiomycota |
| Class: | Agaricomycetes |
| Order: | Boletales |
| Family: | Boletaceae |
| Genus: | Suillellus |
| Species: | S. luridus |
| Binomial name | |
| Suillellus luridus (Schaeff.) Murrill (1909) | |
| Synonyms[1][2] | |
| Suillellus luridus | |
|---|---|
|
| |
| pores on hymenium | |
| cap is convex | |
| hymenium is adnate | |
| stipe is bare | |
| spore print is olive-brown | |
| ecology is mycorrhizal | |
|
edibility: edible but not recommended | |
Suillellus luridus (formerly Boletus luridus), commonly known as the lurid bolete, is a fungus of the bolete family, found in deciduous woodlands on chalky soils in Asia, Europe, and eastern North America. Fruit bodies appear in summer and autumn and may be abundant. It is a solid bolete with an olive-brown cap up to 20 cm (8 in) in diameter, with small red pores on the underside. The stout ochre stem reaches 8–14 cm (3–6 in) high and 1–3 cm (0.4–1.2 in) wide, and is patterned with a red mesh-work. Like several other red-pored boletes, it stains blue when bruised or cut. Though edible when cooked, it can cause gastric upset when eaten raw and can be confused with the poisonous Boletus satanas, though the latter species has a pale cap; as a result, some guidebooks recommend avoiding consumption altogether. When consumed with alcohol, Suillellus luridus has been implicated in causing adverse reactions similar to those caused by the compound coprine, though laboratory testing has not revealed any evidence of coprine in the mushroom.
First described in 1774, the species has been transferred to various Boletaceae genera in its taxonomic history, although it retained the original name given to it by German botanist Jacob Christian Schaeffer until a transfer to genus Suillellus in 2014. Several varieties, a subspecies, and a form have been described by European mycologists. Suillellus luridus is mycorrhizal, forming a symbiotic association with deciduous trees such as oak, birch and beech, and has been found to have a growth-enhancing effect on conifers in experiments. The fruit bodies are highly attractive to, and often infested by, insects, and several species of fly have been recorded feeding on them. Chemical analyses have revealed some aspects of the mushroom's components, including its volatile flavour compounds, its fatty acid and amino acid compositions, and the identities of the carotenoid compounds responsible for its colour.
Taxonomy
Boletus luridus was described by German botanist Jacob Christian Schäffer in 1774, in his series on fungi of Bavaria and the Palatinate, Fungorum qui in Bavaria et Palatinatu circa Ratisbonam nascuntur icones.[3] The specific epithet is the Latin adjective luridus, 'sallow'.[4] French botanist Pierre Bulliard's 1791 Boletus rubeolarius[5] is a heterotypic synonym (based on a different type).[1] The following year, Johann Friedrich Gmelin called it Boletus subvescus,[6] from the Latin words sub "nearly" or "under",[7] and vescus "edible".[8] However, this is a nomen nudum.[9] Several taxonomical synonyms arose when the species was transferred to different genera within the family Boletaceae by different authorities, including Leccinum by Samuel Frederick Gray in 1821,[10] Tubiporus by Petter Karsten in 1881,[11] Dictyopus by Lucien Quélet in 1888,[12] and Suillellus by William Murrill in 1909.[1][13]

The variety Boletus luridus var. erythropus, published as "beta" by Elias Magnus Fries in his 1821 Systema Mycologicum,[14] is synonymous with Boletus erythropus.[15] Boletus luridus var. rubriceps was originally described from Spain (as a species of Tubiporus) by René Maire in 1937,[16] and later formally transferred to Boletus by Aurel Dermek in 1987.[17] Other varieties of B. luridus include Roman Schulz's var. obscurus and var. rubromaculatus published in 1924;[18] Josef Velenovský's 1939 var. tenuipes, found in the Czech Republic;[19] and Jean Blum's 1969 var. lupiniformis and var. queletiformis, originally described from France and Spain, respectively.[20] Boletus erythrentheron, originally described as a distinct species by Jan Bezděk, was later published as the variety B. luridus var. erythrentheron by Albert Pilát and Dermek in 1979,[21] and finally as a subspecies by Jiri Hlavácek in 1995.[22] Carmine Lavorato and Giampaolo Simonini defined the form primulicolor from Sardinia in 1997.[23] Rolf Singer's 1947 variety caucasicus is currently considered an independent species, Boletus caucasicus Singer ex. Alessio.[24] Similarly, B. luridus f. sinensis, found in Hainan Province, China, was later elevated to distinct species status as B. sinensis.[25]
Boletus luridus is the type species of Boletus section Luridi, originally circumscribed by Fries in 1838.[26] This section includes species producing medium to large fruit bodies with thick, swollen stems, and minute pores that are coloured red, orange, or brown.[27] However, the genus as a whole is strongly paraphyletic and will probably be fragmented once further studies have resolved relationships to a finer detail. Manfred Binder and David Hibbett showed B. luridus to be most closely related to a group containing B. torosus and B. luteocupreus, with B. vermiculosus and Pulveroboletus ravenelii as more distant relatives, based on molecular phylogenetics inferred from ribosomal DNA sequences.[28] In a separate molecular study of ten frequently eaten European boletes, B. luridus clustered together with B. rhodoxanthus.[29] Genetic analysis published in 2013 shows that B. luridus and many (but not all) red-pored boletes are part of a dupainii clade (named for Boletus dupainii), well-removed from the core group of Boletus edulis and relatives within the Boletineae.[30] For this reason, it is now considered a species of Suillellus.[31]
The English common name is lurid bolete.[32] Both it and Boletus satanas are known as ayimantari, "bear mushroom", in Eastern Turkey.[33]
Description

Suillellus luridus is a stout fungus with a thick yellow-olive to olive-brown convex cushion-shaped cap that can reach 20 cm (8 in) in diameter.[34] The cap colour tends to darken with age, and regions of red, orange, purple, brown, or olive-green can develop in maturity.[35] The cap surface is tomentose (velvety), becoming smoother with old age, and sticky in wet weather.[36] The pore surface is initially dark red before turning orange-red,[37] and has a lighter-coloured zone encircling the margin.[35] There are 2–3 circular pores per millimetre, and the tubes are 1–2 cm (0.4–0.8 in) deep.[37] The tubes are shorter around the cap margin and close to the stem, where they form a circular depression. Initially pale yellow, the tubes gradually become olive-yellow and then bluish-green upon exposure to air.[35] A characteristic feature is the presence of a maroon layer between the tubes and the flesh.[38] The thick stem is 8–14 cm (3–6 in) tall and 1–3 cm (0.4–1.2 in) wide, and bears a distinctive orange-red meshlike pattern on a paler yellowish or ochre background.[39] The flesh is yellowish, with red marks in the cap, and stains an intense dark blue when bruised or broken. There is a faint sour smell, and the taste is described as mild.[34][36]
Variety queletiformis can be distinguished from the main form by the reddish discolouration of the stem base that occurs both on the exterior surface and in the flesh.[40] Variety rubriceps has a deep crimson red cap, while var. lupiniformis has a pale yellow or dirty ochre cap, sometimes with pink tones throughout.[35]
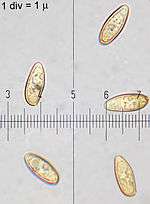
The spore dust is olive to brownish olive. The spores are oval to somewhat fuse-shaped, measuring 11–15 μm long by 4.5–6.5 μm wide.[39] The basidia (spore-bearing cells) are club-shaped and four-spored, and measure 29.2–36.5 by 11.0–12.4 μm. Cystidia on the sides of the tubes (pleurocystidia) are fuse-shaped with swollen middles and long necks, measuring 33–48 by 7.3–13.5 μm; cheilocystidia (on the edges of the pores) have a similar morphology. The cap cuticle is made of cylindrical hyphae 3.7–5.8 μm wide that are interwoven compactly, and the hyphal tips are erect and arranged in bundles. In contrast, the hyphae of the cap flesh is loosely interwoven with hyphae that are cylindrical and branched, measuring 3.7–8.8 μm. Hyphae do not contain clamp connections.[41] The mycelium is an unusual yellow colour.[42]
Some chemical tests can be used to help identify the mushroom. A drop of dilute potassium hydroxide placed on the cap cuticle will stain dark red to blackish, and orange-yellow on the flesh,[37] while ferrous sulphate solution turns the cuticle yellow and then greenish-yellow.[43] Melzer's reagent will turn the flesh dark blue, after the natural bluing reaction to injury has faded.[37]
Similar species
Also found on calcareous (chalky) soils, Rubroboletus satanas is larger and has a pale cap, and its upper stem lacks the reticulations characteristic of S. luridus. Its flesh does not turn blue so markedly on bruising or cutting.[44] It has a more globular overall shape, and young mushrooms carry a smell of decay.[34] The edible Neoboletus luridiformis can be distinguished from S. luridus by its dark brown cap; it also grows on sandy soils associated with conifers.[34] In North America, it can be confused with the poisonous Rubroboletus pulcherrimus, which has a fatter stalk and deeper red pores.[45] Initially collected in Michigan under oak, Boletus vinaceobasis resembles S. luridus but has shorter spores and its cystidia are dark brown in Melzer's reagent.[46] Boletus species that share a similar appearance with S. luridus but lack reticulation in their stems include B. subvelutipes (found in North America and Asia) and B. hypocarycinus (United States).[37]
Boletus caucasicus, found in southern Europe, has yellow flesh in the cap tissue immediately above the tube layer, in contrast to the reddish colour of the same flesh found in S. luridus.[47] Another European species, Rubroboletus rhodoxanthus, has characteristic pinkish tones in the cap.[38] The Chinese species Neoboletus sinensis, originally described as a form of S. luridus, can be most reliably distinguished from the latter by its larger spores, which have been reported to be as large as 12–17 by 5.5–7 μm.[25] Fruit bodies closely resembling S. luridus have been recorded in Australia, though renamed Boletus barragensis as they differ in spore size and preference for trees of the family Myrtaceae.[48][49]
Ecology and distribution
The fungus grows in a mycorrhizal association with deciduous trees such as oak, birch and beech on chalky soils.[34] It is also suspected of being a mycorrhizal associate of subshrub rock roses in the genus Helianthemum.[50] Field studies indicate that the fungus, when paired as a mycorrhizal partner with seedlings of the conifer Cunninghamia lanceolata, increases the seedling's survival rate, augments its height and ground diameter, and increases the chlorophyll content in the leaves.[51] A similar growth-enhancing effect had been noted earlier with Pinus taiwanensis seedlings.[52] These beneficial effects on plant growth are a result of multiple interactions among the fungus, host plant, and indigenous soil microbes that increase the biomass of carbon, and increase the bacterial diversity in the mycorrhizosphere.[53] In a study comparing the salinity resistance of three common ectomycorrhizal fungi (the others were Suillus bovinus and S. luteus), S. luridus was the most tolerant to high concentrations of salt, and is a good candidate species for the inoculation of tree seedlings to be planted on saline soil.[54] Fruit bodies grow singly or scattered on the ground,[37] from June to November after summer rains. S. luridus may occur in parks near a single tree, though it will not be found in acidic soils.[34]

The predilection of insects for this mushroom was noted by 19th-century British mycologist Anna Maria Hussey, who wrote in 1847:
there are very few of the soft-fleshed tribes, all of which are the nurseries of innumerable insects, so much in favour as the poisonous Boletus luridus, on breaking an old one it is a living mass of larvae. Our present subject is so soon attacked by insects that it is very rare to find specimens devoid of wriggling life, and being a very common and abundant kind, it must be of great service in the economy of insect existence.[55]
Several fly species have been recorded feeding on the fruit bodies, including Phaonia boleticola, P. rufipalpis, Thricops diaphanus,[56] and, in North America, Drosophila falleni, Pegomya mallochi, P. winthemi, Megaselia pygmaeoides, and Muscina assimilis.[57] In contrast, slugs tend to avoid consuming this species.[58]
The fungus is widespread in Europe, east to the Black Sea region and eastern Anatolia in Turkey,[33][59] and Pakistan, where it has been recorded from Khanspur, Kuzagali, Sudhan Gali and Hajinpir.[60] In North America, it is known from eastern Canada and the United States east of the Great Lakes, although in one instance it was found in North Dakota.[61] Its range extends south to Mexico,[41][62] and it has been recorded once from Costa Rica.[63] In Asia, it has been found in the Bar'am Forest in the Upper Galilee in northern Israel,[64] and India.[65] Taiwan,[52] It is widely distributed in China, having been found in Hebei, Jiangsu, Anhui, Henan, Guangdong, and Yunnan provinces.[66] The variety rubriceps has been recorded growing under linden (Tilia) in the Czech Republic.[67]
Toxicity
Mild tasting, Suillellus luridus is edible after thorough cooking and is highly regarded in France.[34][44] It is also commonly consumed in Italy and China.[68] The mushroom is sold in regional markets in central Mexico;[69] despite this, it is considered inedible by inhabitants of La Malinche National Park.[62] Caution is advised if choosing this species for consumption, as it resembles other less edible blue-staining boletes. Some guidebooks recommend avoiding it altogether.[4] If eaten raw or insufficiently cooked, symptoms of gastrointestinal poisoning can occur within 30 minutes to two hours, including nausea, vomiting, abdominal cramps, and diarrhoea. A full recovery can be expected within 24 to 48 hours if fluid losses are restored.[70] Chemical analysis has revealed traces of the toxin muscarine and its stereoisomers.[41]
Suillellus luridus has been suspected of causing an enhanced alcohol sensitivity similar to that caused by the common ink cap (Coprinopsis atramentaria), with gastric symptoms. A German mycologist reported having suffered symptoms himself upon imbibing alcohol with this "otherwise excellent" mushroom.[36][71] A 1982 report of three cases from Switzerland further incriminated the species,[72] yet a 1994 study casts doubt on this; researchers Ulrich Kiwitt and Hartmut Laatsch looked for antabuse-like compound coprine content in S. luridus and similar species, and found none in the historical suspect but did find indications for it in the rare B. torosus. They concluded that the most likely explanation for historical incidents was a misidentification of B. torosus with S. luridus, though they could not rule out S. luridus containing a hitherto unidentified compound causing alcohol-related reactions.[73]
Chemistry
The composition of the volatile flavour compounds of Suillellus luridus consists largely of linoleic acid, with smaller proportions of 1-butanol, 3-methyl-1-butanol, pentadecanoic acid, palmitic acid, linoleic acid methyl ester, and heptadecanoic acid. Pyrazine compounds might be responsible for the characteristic odour of the dried mushroom.[74] The predominant sterol present in the fruit bodies is ergosterol, with smaller amounts of closely related derivative compounds.[75] The main fatty acids of the mushroom include linoleic acid (53.4% of total fatty acids), oleic acid (24.1%), and palmitic acid (10.2%). Arginine is the free amino acid found in the highest concentration (96.9 μM per gram of dry weight), followed by glutamine (9.7) and alanine (8.2).[76]
The carotenoid content of the fruit bodies differs substantially between the cap, the tubes, and the stem. The upper part of the cap, which contains 3.1 micrograms of carotenoid per gram (µg/g) fresh weight, has predominantly mutatochrome (47% of total carotenoids), 4-keto-α-carotene (40.2%), and δ-carotene (6.4%). The major carotenoids in the tubes (totaling 4.3 µg/g) include neurosporaxanthin (31.1%), auroxanthin (17.2%), 4-keto-α-carotene (17.1%), and rhodopin (15.8%). The stem (1.2 µg/g) contains primarily auroxanthin (32.5%), followed by 4-keto-α-carotene (19.9%), β-zeacarotene (18.5%), and rhodopin (11.4%).[77] The colour change observed with tissue injury is caused by variegatic and xerocomic acids, both of which turn blue when oxidized enzymatically upon exposure to air.[78]
See also
References
- 1 2 3 "GSD Species Synonymy: Suillellus luridus (Schaeff.) Murrill". Species Fungorum. CAB International. Retrieved 2014-11-24.
- ↑ "Boletus luridus Schaeff., Fungorum qui in Bavaria et Palatinatu circa Ratisbonam nascuntur Icones, 4: 78, t. 107, 1774". MycoBank. International Mycological Association. Retrieved 2013-03-19.
- ↑ Schaeffer JC. (1774). Fungorum qui in Bavaria et Palatinatu circa Ratisbonam nascuntur Icones (in Latin and German). 4. Erlangen: Apud J.J. Palmium. p. 78; plate 107.
- 1 2 Nilson S, Persson O (1977). Fungi of Northern Europe 1: Larger Fungi (Excluding Gill-Fungi). Penguin. p. 104. ISBN 0-14-063005-8.
- ↑ Bulliard JBF. (1791). Herbier de la France (in French). 11. Paris: Chez l'auteur, Didot, Debure, Belin. plate 490.1.
- ↑ Gmelin JF. (1792). Systema Naturae (in Latin). 2 (13th ed.). Leipzig: G.E. Beer. p. 1434.
- ↑ Simpson DP. (1979) [1854]. Cassell's Latin Dictionary (5th ed.). London: Cassell Ltd. pp. 574–75. ISBN 0-304-52257-0.
- ↑ Nilson S, Persson O (1977). Fungi of Northern Europe 2: Gill-Fungi. Penguin. p. 120. ISBN 0-14-063006-6.
- ↑ "Boletus subvescus J.F. Gmel., Systema Naturae, 2: 1434, 1792". MycoBank. International Mycological Association. Retrieved 2013-03-20.
- ↑ Gray SF. (1821). A Natural Arrangement of British Plants. 1. London: Baldwin, Cradock and Joy. p. 648.
- ↑ Karsten PA. (1881). "Enumeratio Boletinearum et Polyporearum Fennicarum, systemate novo dispositarum". Revue Mycologique Toulouse (in Latin). 3: 16–23.
- ↑ Quélet L. (1886). Enchiridion Fungorum in Europa media et praesertim in Gallia Vigentium (in Latin). Lutetia: Octave Dion. p. 160.
- ↑ Murrill WA. (1909). "The Boletaceae of North America – 1". Mycologia. 1 (1): 4–18 (see p. 17). doi:10.2307/3753167.
- ↑ Fries EM. (1821). Systema Mycologicum (in Latin). 1. Lund: Ex Officina Berlingiana. p. 391.
- ↑ "Boletus erythropus Pers., Annalen der Botanik (Usteri), 15: 23, 1795". MycoBank. International Mycological Association. Retrieved 2013-04-17.
- ↑ Marie R. (1937). "Fungi Catalaunici: Series altera. Contributions a l'étude de la flore mycologique de la Catalogne". Publicacions del Instituto Botánico Barcelona (in French). 3 (1): 1–128 (see p. 45).
- ↑ Dermek A. (1987). Boletes III. Fungorum Rariorum Icones Coloratae. 16. Berlin: J. Cramer. pp. 1–23 (see p. 14). ISBN 978-3-443-69002-1.
- ↑ Michael E, Schulz R (1924). Führer für Pilzfreunde: Systematisch geordnet und gänzlich neu bearbeitet von Roman Schulz (in German). 1 (5th ed.). Zwickau: Förster & Borries. plate 92.
- ↑ Velenovský J. (1939). Novitates mycologicae (in Latin). Prague: L. Souc̆ek. p. 158.
- ↑ Petrak F; Commonwealth Mycological Institute (1970). "Index of Fungi". 3 (20): 546. ISSN 0019-3895.
- ↑ Dermek A. (1979). Fungorum rariorum Icones coloratae. 9. J. Cramer. pp. 1–34 (see p. 20). ISBN 978-3-7682-0416-3.
- ↑ Hlavácek J. (1995). "Průzkum naší boletales, 29" [A survey of our Boletales, 29]. Mykologický Sborník (in Czech). 72 (3): 83–90.
- ↑ Lavorato C, Simonini G (1997). "Boletus flavosanguineus sp. nov." (MS word document). Rivista di Micologia (in Italian). 40 (1): 37–51.
- ↑ "Synonymy: Boletus caucasicus Singer ex Alessio, Boletus Dill. ex L. (Saronno): 175 (1985)". Species Fungorum. CAB International. Retrieved 2013-03-20.
- 1 2 Lei Q-Y, Zhou J-J, Wang Q-B (2009). "Notes on three bolete species from China" (PDF). Mycosystema. 28 (1): 56–59. ISSN 1672-6472.
- ↑ Fries EM. (1838). Epicrisis Systematis Mycologici seu Synopsis Hymenomycetum (in Latin). Uppsala: Typographia Academica. p. 417.
- ↑ Snell W, Dick EA (1970). The Boleti of Northeastern North America. Lehre: J. Cramer. p. 79. ISBN 978-0-85486-016-6.
- ↑ Binder M, Hibbett DS (2004). "Part 2: Figure 1C of the Proposed Research". Toward a Global Phylogeny of the Boletales. Clark University. Retrieved 2013-04-13.
- ↑ Mello A, Ghignone S, Vizzini A, Sechi C, Ruiu P, Bonfante P (2006). "ITS primers for the identification of marketable boletes". Journal of Biotechnology. 121 (3): 318–29. doi:10.1016/j.jbiotec.2005.08.022. PMID 16213623.
- ↑ Nuhn ME, Binder M, Taylor AFS, Halling RE, Hibbett DS (2013). "Phylogenetic overview of the Boletineae". Fungal Biology. 117 (7–8): 479–511. doi:10.1016/j.funbio.2013.04.008. PMID 23931115.
- ↑ Vizzini A. (2014). "Nomenclatural novelties" (PDF) (188). Index Fungorum: 1. ISSN 2049-2375.
- ↑ Holden EM. (2003). "Recommended English Names for Fungi in the UK" (PDF). British Mycological Society.
- 1 2 Demirel K, Uzun Y, Kaya A (2004). "Some poisonous fungi of East Anatolia" (PDF). Turkish Journal of Botany. 28 (1/2): 215–19.
- 1 2 3 4 5 6 7 Haas H. (1969). The Young Specialist Looks at Fungi. Burke. p. 126. ISBN 0-222-79409-7.
- 1 2 3 4 Alessio CL. (1985). Boletus Dill. ex L. (sensu lato) (in Italian). Saronno: Biella Giovanna. pp. 169–74.
- 1 2 3 Zeitlmayr L. (1976). Wild Mushrooms: An Illustrated Handbook. Garden City Press, Hertfordshire. p. 102. ISBN 0-584-10324-7.
- 1 2 3 4 5 6 Bessette AR, Bessette A, Roody WC (2000). North American Boletes: A Color Guide to the Fleshy Pored Mushrooms. Syracuse: Syracuse University Press. pp. 126–27. ISBN 0-8156-0588-9.
- 1 2 Michell K. (2006). Field Guide to Mushrooms and Other Fungi of Britain and Europe. London: New Holland Publisher. p. 30. ISBN 1-84537-474-6.
- 1 2 Phillips R. (2006). Mushrooms. London: Pan MacMillan. p. 281. ISBN 0-330-44237-6.
- ↑ Siquier JL, Salom JC (2008). "Contribució al coneixement micologic de les Illes Balears. XVI" (PDF). Revista Catalana de Micologia (in Catalan). 30: 13–25 (see p. 15).
- 1 2 3 Ammirati JA, Traquair JA, Horgen PA (1985). Poisonous Mushrooms of the Northern United States and Canada. Minneapolis: University of Minnesota Press. pp. 237; 240–41. ISBN 0-8166-1407-5.
- ↑ Ramsbottom J. (1953). Mushrooms & Toadstools. Collins. p. 129. ISBN 1-870630-09-2.
- ↑ Dickinson C, Lucas J (1982). VNR Color Dictionary of Mushrooms. Van Nostrand Reinhold. p. 68. ISBN 978-0-442-21998-7.
- 1 2 Lamaison J-L, Polese J-M (2005). The Great Encyclopedia of Mushrooms. Cologne: Könemann. p. 31. ISBN 3-8331-1239-5.
- ↑ Arora D. (1986). Mushrooms Demystified: A Comprehensive Guide to the Fleshy Fungi. Berkeley: Ten Speed Press. p. 528. ISBN 0-89815-169-4.
- ↑ Smith AH, Thiers HD (1971). The Boletes of Michigan. Ann Arbor: University of Michigan Press. pp. 347–48. ISBN 0-472-85590-5.
- ↑ Loizides M. (2011). "Quercus alnifolia: The indigenous Golden Oak of Cyprus and its fungi". Field Mycology. 12 (3): 81–88. doi:10.1016/j.fldmyc.2011.06.004.
- ↑ Watling R, Li TH (1999). Australian Boletes: A Preliminary Survey. Royal Botanic Garden Edinburgh. p. 33. ISBN 1-872291-28-7.
- ↑ Grgurinovic CA. (1997). Larger Fungi of Southern Australia. Adelaide: Botanic Gardens of Adelaide & State Herbarium. p. 229. ISBN 978-0-7308-0737-7.
- ↑ Barden N. (2007). "Helianthemum grasslands of the Peak District and their possible mycorrhizal associates". Field Mycology. 8 (4): 119–26. doi:10.1016/S1468-1641(10)60384-2.
- ↑ Wang Y-J, Tang M, Guo Y, Zhang F-F, Huang Y-H (2006). 外生菌根真菌对杉木的接种效应 [Inoculation effect of ectomycorrhizal fungi on Cunninghamia lanceolata]. Acta Botanica Boreali-Occidentalia Sinica (in Chinese and English). 26 (9): 1900–04. ISSN 1000-4025.
- 1 2 Hu H-T. (1981). "Mycorrhizae of some important tree species grown at high elevation in Taiwan". National Science Council Monthly (in Chinese). 9 (4): 303–34. ISSN 0250-1651.
- ↑ Zhang H-H, Tang M, Chen H, Zheng C-L (2010). "Effects of inoculation with ectomycorrhizal fungi on microbial biomass and bacterial functional diversity in the rhizosphere of Pinus tabulaeformis seedlings". European Journal of Soil Biology. 46 (1): 55–61. doi:10.1016/j.ejsobi.2009.10.005.
- ↑ Sheng M, Chen H, Zhang FF (2009). "In vitro salinity resistance of three ectomycorrhizal fungi". Soil Biology and Biochemistry. 41 (5): 948–53. doi:10.1016/j.soilbio.2008.12.007.
- ↑ Hussey AM. (1847). Illustrations of British Mycology, Containing the Figures and Description of Funguses of Interest and Novelty Indigenous to Britain. London: Reeve, Brothers, King William Street, Strand. Description of plate XXIII.
- ↑ Skidmore P. (1985). The Biology of the Muscidae of the World. Dordrecht & Boston: Springer. pp. 159, 493. ISBN 978-90-6193-139-3.
- ↑ Bruns T. (1984). "Insect mycophagy in the Boletales: Fungivore diversity and the mushroom habitat". In Wheeler Q, Blackwell M. Fungus-Insect Relationships: Perspectives in Ecology and Evolution. New York: Columbia University Press. pp. 91–129. ISBN 0-231-05694-X.
- ↑ Elliott WT. (1922). "Some observations on the mycophagous propensities of slugs". Transactions of the British Mycological Society. 8 (1–2): 84–90. doi:10.1016/S0007-1536(22)80011-5.
- ↑ Sesli E. (2007). "Preliminary checklist of macromycetes of the East and Middle Black Sea Regions of Turkey" (PDF). Mycotaxon. 99: 71–74.
- ↑ Sarwar S, Khalid AN (2013). "Preliminary Checklist of Boletales in Pakistan" (PDF). Mycotaxon: 1–12.
- ↑ Stack RW. (1981). "Rare mycorrhizal fungi fruiting in North Dakota in 1980". Proceedings of the North Dakota Academy of Science. 35: 44. ISSN 0096-9214.
- 1 2 Montoya A, Kong A, Estrada-Torres A, Cifuentes J, Caballero J (2004). "Useful wild fungi of La Malinche National Park, Mexico" (PDF). Fungal Diversity. 17: 115–43.
- ↑ Halling RE, Mueller GM (2008). "Boletus luridus". New York Botanic Garden: Macrofungi of Costa Rica. New York Botanic Garden. Retrieved 2008-02-04.
- ↑ Avizohar-Hershenzon Z, Binyamini N (1972). "Boletaceae of Israel: I. Boletus sect. Luridi". Transactions of the British Mycological Society. 59 (1): 25–30. doi:10.1016/s0007-1536(72)80037-8.
- ↑ Kumar S, Sharma YP (2011). "Additions to boletes from Jammu and Kashmir". Journal of Mycology and Plant Pathology. 41 (4): 579–83.
- ↑ Zhishu B, Zheng G, Taihui L (1993). The Macrofungus Flora of China's Guangdong Province (Chinese University Press). New York: Columbia University Press. p. 487. ISBN 962-201-556-5.
- ↑ Tichy H. (1998). "Hřib koloděj červenohlavý Boletus luridus var. rubriceps (R. Maire) roste v parku" [Boletus luridus var. rubriceps (R. Maire) growing in a park]. Mykologicky Sbornik (in Czech and English). 75 (1): 20–21. ISSN 0374-9436.
- ↑ Rubel W, Arora D (2008). "A study of cultural bias in field guide determinations of mushroom edibility using the iconic mushroom, Amanita muscaria, as an example". Economic Botany. 63 (3): 223–43. doi:10.1007/s12231-008-9040-9.
- ↑ Pérez-Moreno J, Martínez-Reyes M, Yescas-Pérez A, Delgado-Alvarado A, Xoconostle-Cázares B (2008). "Wild mushroom markets in central Mexico and a case study at Ozumba". Economic Botany. 62 (3): 425–36. doi:10.1007/s12231-008-9043-6.
- ↑ Tomalak M, Rossi E, Ferrini F, Moro PA (2011). "Negative aspects and hazardous effects of forest environment on human health". In Nilsson K, et al. Forests, Trees and Human Health. New York: Springer. pp. 77–124 (see p. 101). ISBN 978-90-481-9805-4.
- ↑ Flammer R. (1985). "Brechdurchfalle als Leitsymptom – Pilze und Alkohol" [Diarrhoea and vomiting as main symptoms – mushrooms and alcohol]. Praxis (in German). 74 (37): 992–96.
- ↑ Benjamin DR. (1995). Mushrooms: Poisons and Panaceas—A Handbook for Naturalists, Mycologists and Physicians. New York: WH Freeman and Company. p. 291. ISBN 0-7167-2649-1.
- ↑ Kiwitt U, Laatsch H (1994). "Coprin in Boletus torosus: Beruht die angebliche Alkoholunverträglichkeit durch den Verzehr des Netzstieligen Hexenröhrlings (Boletus luridus) auf einer Verwechslung?" [Coprine in Boletus torosus: Is the alleged alcohol hypersensitivity by ingestion of B. luridus caused by a mistake?] (PDF). Zeitschrift für Mykologie (in German). 60 (2): 423–30.
- ↑ Li W-W, Wu S-M, Xu T (2009). 褐黄牛肝菌挥发性风味物质组成研究 [Volatile flavor composition of Boletus luridus Schaeff.: Fr.]. Journal of Shanghai Jiaotong University – Agricultural Science (in Chinese and English). 27 (3): 300–04. doi:10.3969/j.issn.1671-9964.2009.03.022.
- ↑ De Simone F, Senatore F, Sica D, Zollo F (1979). "Sterols from some basidiomycetes". Phytochemistry. 18 (9): 1572–73. doi:10.1016/S0031-9422(00)98504-2.
- ↑ Dembitsky VM, Terent'ev AO, Levitsky DO (2010). "Amino and fatty acids of wild edible mushrooms of the genus Boletus". Records of Natural Products. 4 (4): 218–23. ISSN 1307-6167.
- ↑ Czezuga B. (1978). "Investigations on carotenoids in fungi IV. Members of the Boletus genus". Qualitas Plantarum. 28 (1): 37–43. doi:10.1007/BF01092999.
- ↑ Nelson SF. (2010). "Bluing components and other pigments of Boletes" (PDF). FUNGI. 3 (4): 11–14.
{kind=link}
{kind=link}
External links
| Wikimedia Commons has media related to Suillellus luridus. |
 Data related to Suillellus luridus at Wikispecies
Data related to Suillellus luridus at Wikispecies- Suillellus luridus in Index Fungorum