Protein O-GlcNAcase
| Protein O-GlcNAcase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifiers | |||||||||
| EC number | 3.2.1.169 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Protein O-GlcNAcase (EC 3.2.1.169, OGA, glycoside hydrolase O-GlcNAcase, O-GlcNAcase, BtGH84, O-GlcNAc hydrolase) is an enzyme with systematic name (protein)-3-O-(N-acetyl-D-glucosaminyl)-L-serine/threonine N-acetylglucosaminyl hydrolase.[1][2][3][4][5] This enzyme catalyses the following chemical reaction
- (1) [protein]-3-O-(N-acetyl-beta-D-glucosaminyl)-L-serine + H2O [protein]-L-serine + N-acetyl-D-glucosamine
- (2) [protein]-3-O-(N-acetyl-beta-D-glucosaminyl)-L-threonine + H2O [protein]-L-threonine + N-acetyl-D-glucosamine

Isoforms
There are three isoforms of O-GlcNAcase in humans that have been identified. Full-length O-GlcNAcase (fOGA), the shortest O-GlcNAcase (sOGA), and a variant of OGA (vOGA). The human OGA gene is capable of producing two separate transcriptions, each capable of encoding a different OGA isoform. The long isoform gene codes for fOGA, a bifunctional enzyme that primarily resides in the cytoplasm. In contrast, vOGA resides within the nucleus. However, all three isoforms exhibit glycoside hydrolase activity.[6]
Homologs
Homologs exist in other species as O-GlcNAcase is conserved in higher eukaryotic species. In a pairwise alignment, humans share 55% homology with Drosophilia and 43% with C. elegans. Drosophilia and C. elegans share 43% homology. Among mammals, the OGA sequence is even more highly conserved. The mouse and the human have 97.8% homology. However, OGA does not share significant homology with other proteins. However, short stretches of ~200 amino acids in OGA have homology with some proteins such as hyaluronidase, a putative acetyltransferase , eukaryotic translation elongation factor-1γ, and the 11-1 polypeptide. [7]
Reaction
Protein O-GlcNAcylation
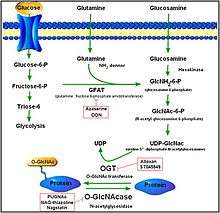
O-GlcNAcylation is a form of glycosylation, the site-specific enzymatic addition of saccharides to proteins and lipids. This form of glycosylation is with O-linked β-N-acetylglucosamine or beta-O-linked 2-acetamido-2-deoxy-d-glycopyranose (O-GlcNAc). In this form, a single sugar (β-N-acetylglucosamine) is added to serine and threonine residues of nuclear or cytoplasmic proteins. Two conserved enzymes control this glycosylation of serine and threonine: O-GlcNAc transferase (OGT) and O-GlcNAcase (OGA). While OGT catalyzes the addition of O-GlcNAc to serine and threonine, OGA catalyzes the hydrolytic cleavage of O-GlcNAc from post-transitionally modified proteins.[8]
OGA is a member of the family of hexosaminidases. However, unlike lysosomal hexosaminidases, OGA activity is the highest at neutral pH (~7) and it localizes mainly to the cytosol. OGA and OGT are synthesized from two conserved genes (OGA is encoded by MGEA5) and are expressed throughout the human body with high levels in the brain and pancreas. The products of O-GlcNAc and the process itself plays a role in embryonic development, brain activity, hormone production, and a myriad of other activities. [9] [10]
Over 600 proteins are targets for O-GlcNAcylation. While the functional effects of O-GlcNAc modification is not fully known, it is known that O-GlcNAc modification impacts many cellular activities such as lipid/carbohydrate metabolism and hexosamine biosynthesis. Modified proteins may modulate various downstream signaling pathways by influencing transcription and proteomic activities. [11]
Regulation
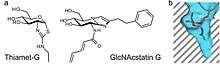
OGA catalyzes O-GlcNAc hydrolysis via an oxazoline reaction intermediate. Stable compounds which mimic the reaction intermediate can act as selective enzyme inhibitors. Thiazoline derivatives of GlcNAc can be used as a reaction intermediate. An example of this includes Thiamet-G as shown on the right. A second form of inhibition can occur from the mimicry of the transition state. The GlcNAcstatin family of inhibitors exploit this mechanism in order to inhibit OGA activity. For both types of inhibitors, OGA can be selected apart from the generic lysosomal hexosaminidases by elongating the C2 substituent in their chemical structure. This takes advantage of a deep pocket in OGA's active site that allow it to detect analogs of GlcNAc. [12]
Regulation of O-GlcNAcase can be taken advantage of in various medical procedures. One such example is the use of the aforementioned Thiamet-G in Alzheimer's disease treatment. When the tau protein in the brain is hyperphosphorylated, neurofibrillary tangles form, which are a pathological hallmark for neurodegenerative diseases such as Alzheimer's disease. In order to treat this condition, OGA is targeted by inhibitors such as Thiamet-G in order to prevent O-GlcNAc from being removed from tau, which assists in preventing tau from becoming phosphorylated. [13]
References
- ↑ Wells, L.; Gao, Y.; Mahoney, J.A.; Vosseller, K.; Chen, C.; Rosen, A.; Hart, G.W. (2002). "Dynamic O-glycosylation of nuclear and cytosolic proteins: further characterization of the nucleocytoplasmic β-N-acetylglucosaminidase, O-GlcNAcase". J. Biol. Chem. 277 (3): 1755–1761. doi:10.1074/jbc.M109656200. PMID 11788610.
- ↑ Cetinbas, N.; Macauley, M.S.; Stubbs, K.A.; Drapala, R.; Vocadlo, D.J. (2006). "Identification of Asp174 and Asp175 as the key catalytic residues of human O-GlcNAcase by functional analysis of site-directed mutants". Biochemistry. 45 (11): 3835–3844. doi:10.1021/bi052370b. PMID 16533067.
- ↑ Dennis, R.J.; Taylor, E.J.; Macauley, M.S.; Stubbs, K.A.; Turkenburg, J.P.; Hart, S.J.; Black, G.N.; Vocadlo, D.J.; Davies, G.J. (2006). "Structure and mechanism of a bacterial β-glucosaminidase having O-GlcNAcase activity". Nat. Struct. Mol. Biol. 13 (4): 365–371. doi:10.1038/nsmb1079. PMID 16565725.
- ↑ Kim, E.J.; Kang, D.O.; Love, D.C.; Hanover, J.A. (2006). "Enzymatic characterization of O-GlcNAcase isoforms using a fluorogenic GlcNAc substrate". Carbohydr. Res. 341 (8): 971–982. doi:10.1016/j.carres.2006.03.004. PMID 16584714.
- ↑ Dong, D.L.; Hart, G.W. (1994). "Purification and characterization of an O-GlcNAc selective N-acetyl-β-D-glucosaminidase from rat spleen cytosol". J. Biol. Chem. 269 (30): 19321–19330. PMID 8034696.
- ↑ Li, J.; Huang, C.L.; Zhang, L.W.; Lin, L.; Li, Z.H.; Zhang F.W; Wang P. (2010). "Isoforms of human O-GlcNAcase show distinct catalytic efficiencies". Biochemistry (Mosc.). 75: 938–943. doi:10.1134/S0006297910070175. PMID 20673219.
- ↑ Gao, Y.; Wells, L.; Comer, F.I.; Parker, G.J.; Hart, G.W. (2001). "Dynamic O-glycosylation of nuclear and cytosolic proteins: cloning and characterization of a neutral, cytosolic β-N-acetylglucosaminidase from human brain". J. Biol. Chem. 276: 9838–9845. doi:10.1074/jbc.M010420200. PMID 11148210.
- ↑ Lima, V.; Rigsby, C.; Hardy, D.; Webb, R.; Tostes, R. (2009). "O-GlcNAcylation: a novel post-translational mechanism to alter vascular cellular signaling in health and disease: focus on hypertension". Journal of the American Society of Hypertension. 3: 374–387. doi:10.1016/j.jash.2009.09.004.
- ↑ Förster, S.; Welleford, A.; Triplett, J.; Sultana, R.; Schmitz, B.; Butterfield, D. (2014). "Increased O-GlcNAc Levels Correlate with Decreased O-GlcNAcase Levels in Alzheimer Disease Brain". Biochim Biophys Acta. 1842: 1333–1339. doi:10.1016/j.bbadis.2014.05.014.
- ↑ Shafi, R.; Iyer, SP.; Ellies, L.; O'Donnell, N.; Marek, KW.; Chui, D.; Hart, GW.; Marth, JD. (2000). "The O-GlcNAc transferase gene resides on the X chromosome and is essential for embryonic stem cell viability and mouse ontogeny". Proc Natl Acad Sci U S A. 97: 5735–5739. doi:10.1073/pnas.100471497. PMID 10801981.
- ↑ Love, D.; Ghosh, S.; Mondoux, M.; Fukushige, T.; Wang, P.; Wilson, M.; Iser, W.; Wolkow, C.; Krause, M.; Hanover, J. (2010). "Dynamic O-GlcNAc cycling at promoters of Caenorhabditis elegans genes regulating longevity, stress, and immunity". Proc Natl Acad Sci U S A. 107: 7413–7418. doi:10.1073/pnas.0911857107.
- ↑ Alonso, J.; Schimpl, M.; Aalten, D. (2014). "O-GlcNAcase: Promiscuous Hexosaminidase or Key Regulator of O-GlcNAc Signaling?". The Journal of Biological Chemistry. 289: 34433–34439. doi:10.1074/jbc.R114.609198.
- ↑ Lim, S.; Haque, M.; Nam, G.; Ryoo, N; Kim, Y. (2015). "Monitoring of Intracellular Tau Aggregation Regulated by OGA/OGT Inhibitors". International Journal of Molecular Sciences. 16: 20212–20224. doi:10.3390/ijms160920212. PMID 26343633.
External links
- Protein O-GlcNAcase at the US National Library of Medicine Medical Subject Headings (MeSH)